Лесные плантации — наиболее продуктивные лесные массивы. Они дают древесины с единицы площади в 2—2,5 раза больше, чем неокультуренные равнинные леса; мало отличаются от сельскохозяйственных угодий — их обрабатывают средствами защиты, удобряют, вырубают в строгом соответствии с природным циклом развития. На эти леса незначительно влияет закисление почв в результате выпадения кислотных дождей. Однако они подвержены воздействию кислотных дождей через хвою или листву. При этом большое значение имеет их географическое расположение (так, в прибрежных и в высокогорных районах главным фактором повреждений могут быть туманы).[ ...]
Окультуренные леса характеризуются средней продуктивностью. Они частично восполняются за счет новых посадок, их обрабатывают средствами против вредителей, защищают от пожаров, прореживают и известкуют.[ ...]
Неокультуренные равнинные леса во многих странах занимают большую территорию. Они располагаются на территориях национальных парков, водоохранных зон, заказников, лесоводческих хозяйств. Их окультуривают только в небольшой "степени (в основном для защиты от пожаров).[ ...]
С августа по сентябрь среднесуточная концентрация озона составляет 80—110 б.д. с максимумами 190—250 б.д. при средней продолжительности таких эпизодов 2 ч. При этом наблюдается интересное явление. В отличие от городов в лесном массиве концентрация озона в ночное время практически не уменьшается (80 б.д.). В городах в ночное время в результате реакций озона с оксидами азота и летучими органическими соединениями концентрация обычно снижается весьма значительно (не выше 60 б.д.).[ ...]
Симптомы болезни леса стали появляться у сосны с конца 50-х годов — времени возникновения фотохимического смога в Калифорнии. Замедление роста хвойных пород в довольно отдаленных от Лос-Анджелеса зонах (в Секвойе и Королевском каньоне Национального парка) началось с 1965 г. Среднесуточная концентрация озона составила 60—85 б.д. с максимумами дб 160 б.д. Для выяснения роли озона в гибели деревьев в 1962 г. были проведены полевые испытания. Молодые ели помещали в полиэтиленовые мешки и в течение 9—18 сут. ежедневно обрабатывали 9 ч озоном (500 б.д.). В результате этого у них появились симптомы, характерные для поврежденного леса,— усыхание вершин деревьев (основной признак), хлороз и мозаичная пятнистость хвои, опадание иголок.[ ...]
Красная ель. Эта хвойная порода представляет интерес с точки зрения гибели лесов в высокогорных зонах. Заметное ухудшение состояния этих лесов было отмечено в начале 50-х годов, причем симптомы были особенно сильно выражены у елей, растущих на высоте 1000—1500 м над уровнем моря. Красная ель произрастает на востоке США, в северных Аппалачах, часто соседствует с бальзамической пихтой и пихтой Фрезера. Периодические наблюдения за лесами, где отмечена гибель красной ели, позволили установить механизм повреждений хвойных пород. В высокогорных зонах кроны деревьев находятся в облачном слое до 3000 ч/год, pH дождей равен 3,6. pH облаков, как правило, на 0,5 ед. ниже, чем дождей (т. е. они более кислые). В облаках отмечена высокая концентрация пероксида водорода (в среднем 1000 б.д. в летний период). Почвы, на которых произрастает красная ель, довольно бедные и содержат большое количество токсичных металлов.[ ...]
У красной ели при действии кислых облаков в первую очередь повреждаются молодые побеги, внешне это проявляется в усыхании кроны от верхушки вниз и от наружной части веток внутрь. Этот процесс принципиально отличается от повреждений хвойных пород, произрастающих на равнинах: прежде всего повреждаются старые иглы и побеги, поэтому дефолиация идет снизу вверх и от внутренней части веток наружу. Наиболее интенсивно повреждаются ели, находящиеся в облаках в летний период более половины времени.[ ...]
Чтобы установить влияние структуры почвы на повреждаемость красной ели при постоянном уровне кислотности тумана (500—1000 ч/год, pH 3,25) и концентрации озона ( 80 б.д), в сопоставимых условиях определяли органолептические показатели и инкременты годового радиального роста в зависимости от толщины почвенного слоя. Наименьшая скорость роста (0,9 мм/год) и повреждение хвои (хлороз, прогрессирующее опадание хвои, сокращение продолжительности развития новых побегов) отмечены на тонких слоях почвы (20—25 см на гранитной основе), при более значительной толщине почвенного слоя инкремент радиального роста составил 1,6 мм/год; на глубоких черноземных слоях изменений хвои не отмечено и скорость роста максимальна (2,5 мм/год). Исходя из изложенного, причинами повреждения красной ели можно считать истощение почв и нарушение обмена веществ под действием озона и кислого тумана.[ ...]
Сахарный клен. Наиболее систематические исследования лиственных лесов проведены в США и Канаде на посадках сахарного клена. С начала 70-х годов наблюдаются прогрессирующие повреждения крон, которые проявляются в дефолиации, уменьшении площади листа, хлорозе, усыхании вершин деревьев, отслаивании коры. При этом резко падает продуктивность сокоотделения. Установлено, что из общей площади посадок в провинции Квебек (Канада) у 10 % деревьев полностью усохли вершины стволов, в 35 % — средние повреждения (11—25% дефолиации), 4% сильно повреждены (25—50 % дефолиации), 1%—с дефолиацией более 50 %. Листва опадает максимально в конце лета, в результате чего снижается резервный запас углеводов и замедляется развитие в следующем году. Наиболее заметно это проявляется на бедных почвах. На больных деревьях интенсивно размножаются вредители: кленовый мотылек и гусеницы. Повреждения кленовых массивов обусловлены как влиянием озона и кислотных дождей на крону, так и изменениями в составе почвы. С 1977 г. на 75 % территории провинции Квебек, занятой кленовыми посадками, значительно уменьшилось содержание в почве ионов К+, Са2 + и М§2+ как в верхнем горизонте (гумусовый слой), так и в нижнем.[ ...]
Хвойные и лиственные леса в центральной Европе. Гибель лесов прогрессирует с 70-х годов по настоящее время.[ ...]
Рисунки к данной главе:
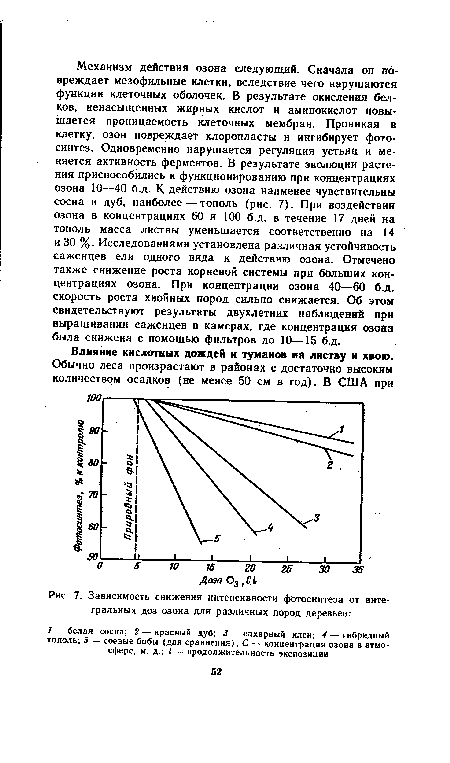
| Зависимость снижения интенсивности фотосинтеза от интегральных доз озона для различных пород деревьев |
 |