В опытах, поставленных нами на кафедре, выделение Н-ионов корнями пшеницы и риса в обмен н адсорбируемые ими катионы из ионитов в песчаных культурах было значительно выше, чем в солевой питательный раствор. Это свидетельствует, что усвоение корнями ионов из раствора требует меньших «затрат» со стороны растения по сравнению с поглощением тех же ионов из адсорбированного сосфяния. Вместе с тем дальнейшие эксперименты позволили установить, чтф обменный Н-фонд растения зависит от уровня обмена веществ в организме: при интенсивном обмене веществ усиливается и поглощение ионов извне и выделение водородных ионов во внешнюю среду.[ ...]
Некоторые закономерности поступления питательных веществ в корневую систему можно проверить и на простых, модельных опытах. Если взять, например, мешочек из целлюлозы, заполнить его ионитами (насыщенными катионами Н’ и анионами НСО , то есть так же, как это бывает и в случае корневых волосков) и ввести во влажную почву, содержащую необходимые растению питательные соли, то спустя некоторое время (этот искусственный «корень» погружают в почву на 1—2 суток) можно обнаружить обмен ионами между содержимым мешочка и почвой. Не представляет труда вытеснить эти ионы из ионитов и определить количественно.[ ...]
Подобно корневым войоскам, эта модель может извлечь из наружного раствора ионы довольно Полно, так как равновесие не устанавливается: все адсорбированные ионы удаляются из раствора, что способствует течению реакции обмена между «корнем» и внешней средой.[ ...]
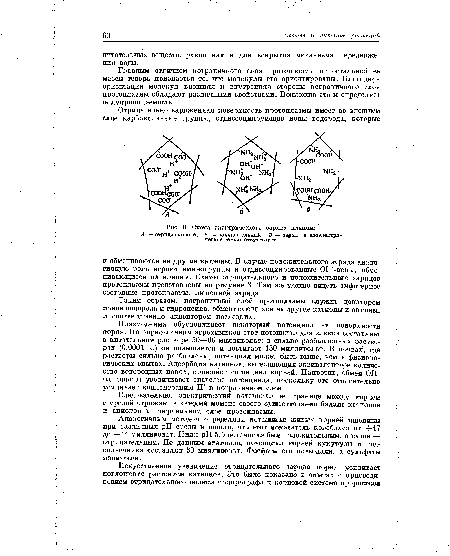
Главным отличием пограничного слоя протоплазмы от остальной ее массы теперь признается то, что молекулы его ориентированы. Благодаря ориентации молекул внешняя и внутренняя стороны пограничного слоя протоплазмы обладают различными свойствами. Возможно это и определяет полупроницаемость.[ ...]
А — отрицательный; Б — положительный; В — заряа в изоэлектри-ческой точке отсутствует.[ ...]
Таким образом, пограничный слой протоплазмы служит донатором ионов водорода и гидроксила, обменивающихся на другие катионы и анионы, и соответственно акцептором последних.[ ...]
Плазмолемма обусловливает некоторый потенциал на поверхности корня. По определениям агрохимиков этот потенциал для злаков составляет в питательном растворе 50—60 милливольт; в сильно разбавленных растворах (0,0001 н.) он повышается и достигает 150 милливольт. В почвах, где растворы сильно разбавлены, потенциал может быть выше, чем в физиологических опытах. Адсорбция катионов, вытесняющих эквивалентное количество водородных ионов, понижает потенциал корней. Напротив, обмен ОН на анионы увеличивает значение потенциала, поскольку это относительно усиливает концентрацию Н в пограничном слое.[ ...]
Следовательно, электрический потенциал на границе между корнем и средой отражает в каждый момент своего существования баланс катионов и анионов в пограничном слое протоплазмы.[ ...]
Аналогичным методом определяли потенциал живых корней пшеницы при различных pH среды и нашли, что этот показатель колебался от +13 до —11 милливольт. Ниже pH 5,5 потенциал был положительным, а выше — отрицательным. По данным анализов, потенциал корней кукурузы и подсолнечника составлял 30 милливольт. Фосфаты его повышали, а сульфаты понижали.[ ...]
Рисунки к данной главе:
| Схема электрического заряда плазмы |
 |
Аналогичные главы в дргуих документах:
| См. далее:Поверхностный слой протоплазмы и его роль в поглощении питательных веществ корневой системой |