Биогеографические, или эколого-географические, закономерности довольно сложно классифицировать. Как только что упомянуто, любые явления происходят в пространстве, а его протяженность не может быть математически определена в формальных единицах «от сих до сих», поскольку в экологии речь идет о системной величине. Для микроорганизма кончик иглы уже огромный полигон. К тому же в обширной биогеографи-ческой литературе широко освещены весьма значительные эмпирические данные. Существуют многочисленные сводки по географии растений, животных, общей биогеографии, географии сообществ, биогеографии островов и так далее. Приводимые факты как-то обобщены, сформулировано множество частных закономерностей. Из этой массы ценных данных здесь сделана попытка выделить наиболее общие принципы, объединяемые в три основные группы: ареал и распространение видов в его пределах, изменения особей (популяций) в пределах ареала и закономерности распространения сообществ. Многие смежные вопросы, обычно также относимые к биогеографическим, рассмотрены в следующем разделе 3.8.[ ...]
Если в какой-то географической точке никто никогда не вел фаунисти-ческие или флористические исследования, точек находок, естественно, не возникает. Кроме того, при многолетнем изучении широких регионов, как это приходилось делать автору в Средней Сибири, нетрудно убедиться, что некоторые птицы, например, коростель, порой фактически исчезают на краю своего ареала на ряд лет, а затем вновь появляются. Для упомянутого вида территориальная изменчивость границ ареала составляет сотни километров, так что выразима в самом мелком масштабе. Для менее подвижных животных и растений такие колебания границ ареала невозможны, но в годы депрессий численности экологическая роль видов, находящихся в минимуме, практически равна нулю или стремится к нему. Исследователь может вид не обнаружить по чисто техническим причинам.[ ...]
Возможности сопряжения и прогнозирования границ ареалов связаны с конгруэнтностью видов в составе биоценозов и сообществ (разд. 3.8.). Как показывает опыт, имеется соответствие видов и ценозов (см. ниже), и эти физиономически ясные черты позволяют отображать такие закономерности на картах. Однако довольно трудно (хотя и возможно) найти два вида, которые бы имели абсолютно сходное распределение в пространстве. Эту особенность Л. Г. Раменский (1924) и Г. А. Глизон (1926) сформулировали в виде индивидуалистической гипотезы, правила или принципа экологической индивидуальности видов: каждый вид пространственно распределен в соответствии со своими генетическими, физиологическими и другими биологическими особенностями и глубоко специфично относится к факторам среды, в том числе к другим видам. Как нет абсолютно идентичных особей (кроме однояйцевых близнецов, тем не менее нередко фенотипически отличающихся друг от друга), так нет и биоэкологически идентичных видов. Следовательно, их распространение сугубо индивидуально, а потому подвержено столь же индивидуальным нормам реакции на возникающие изменения среды жизни. Как сказано выше, этот принцип не абсолютен.[ ...]
Как показывают широкомасштабные исследования в Сибири и на Дальнем Востоке, виды очень четко соответствуют ценозам, а если наблюдается разница в постоянно регистрируемом видовом составе всего в один вид, то ценоз явно резко отличается от географически или топографически соседних.[ ...]
Как правило, смена видового состава происходит не на один, а на группу видов, и действительно, вид служит индикатором того сообщества, куда он входит.[ ...]
Необходимо обратить внимание на то, что в ходе упомянутых процессов нарушается принцип территориальной общности физико-географических единиц (разд. 3.7.3), они укрупняются из-за мозаичности возникающих «обрывков» экосистем и их возрастающей отдаленности друг от друга. Иногда «промежуточные» экосистемы (по иерархии соподчинения) вообще исчезают, как, впрочем, и элементарные, в своем первозданном виде. Иерархия экосистем резко упрощается по числу уровней — сохраняется как целое лишь 2-й снизу уровень биома или ландшафтной полосы (см. главу 2) и только им соответствуют по распространению виды, занимая микроучастки, едва ли составляющие полноценные экосистемы (скорее, их парацеллы).[ ...]
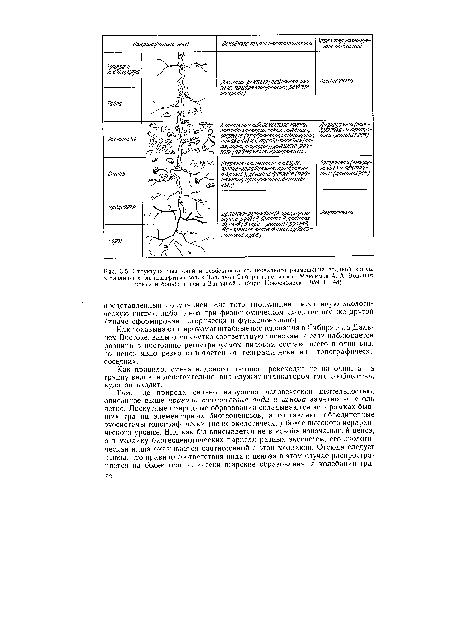
Там, где сохранилась первичная структура природных образований, соответствие вида и ценоза дает очень четкую пространственную картину кружева ареала, закономерно меняющегося при движении в географическом пространстве. Эта закономерность в разд. 3.6 была названа правилом топографического, или популяционного, кружева ареала. Это правило было сформулировано для топографического пространства одной географической точки. Теперь следует обратить внимание на изменения этого кружева в более широком пространстве — правило географической изменчивости кружева ареала. Обычно оно состоит в том, что на севере вид приурочен к приречным, низменным участкам, затем к югу заселяет более широкие пространства, в горах обитает лишь в локальных участках, а на южном пределе распространения вновь приурочен к приречным более влажным и прохладным местообитаниям. У каждого широко распространенного вида характер изменения кружева ареала специфичен, но смены происходят закономерно. Такие смены, пожалуй, лучше всего известны для водяной крысы в Сибири1 (рис. 3.5). Пользуясь выявленными закономерностями, можно получить более надежные критерии для экстраполяции границ ареалов видов.[ ...]
Проверка приведенных теоретических положений в свое время была сделана автором на Дальнем Востоке методом артиллерийской «вилки», т. е. были намечены точки, где вид уже не наблюдался и где наверняка существовал, а в промежуточном регионе по характеру растительности части экосистемы должен был бы присутствовать. В этой срединной точке проводились исследования. Все ожидавшиеся виды (в том числе не склонные к флуктуации границ ареала) были обнаружены вне зависимости от того, на каком расстоянии от точки наблюдения были проведены на картах ранее известные предельные границы их распространения. Часто находка была в сотнях километров к северу от «северного предела распространения» вида.[ ...]
Наиболее общим объяснением причин формирования границ ареала вида служит правило ограничивающих факторов: факторы среды, наиболее удаляющиеся от оптимума экологических потребностей вида, лимитируют возможности его существования в данных условиях. Поскольку к лимитирующим факторам относятся любые условия существования вида — как абиотические, так и биотические, включая антропогенные,— правило ограничивающих факторов, ведущее свое начало от группы законов минимума (см. разд. 3.5.2), включая закон толерантности Шел-форда (см. разд. 3.5.1), практически дополнительно ничего не объясняет, а лишь резюмирует перечисленные закономерности.[ ...]
Для объяснения механизма действия принципа воздействия факторов В. Тишлера (см. выше) было предложено несколько теоретических оснований. Одно из них — правило стимулирующего действия температур В. Шелфорда — Т. Парка, в умеренных широтах определяющее стимулирующее воздействие, которое оказывают изменения температуры среды обитания (правило сформулировано независимо друг от друга В. Э. Шел-фордом в 1929 г. и Т. Парком в 1930 г.). Это правило в 1972 г. А. Н. Голиков и О. А. Скарлато дополнили правилом лимитирующего значения крайних летних и зимних температур, или «биогеографическим методом» указанных авторов: в умеренных широтах Северного полушария распространение видов ограничено необходимыми для размножения температурами. Эта потребность является генотипически закрепленным видовым признаком и прямо не связана с диапазоном температур выживаемости особей вида. У видов, переносящих широкие колебания температур, температура размножения часто более контрастна, чем у видов с относительно низким порогом выносливости к переменам количества тепла.[ ...]
Рисунки к данной главе:
| Структура поселений и особенности стационарного размещения водяной крысы в различных ландшафтных зонах Западной Сибири (рис. из кн. |
 |