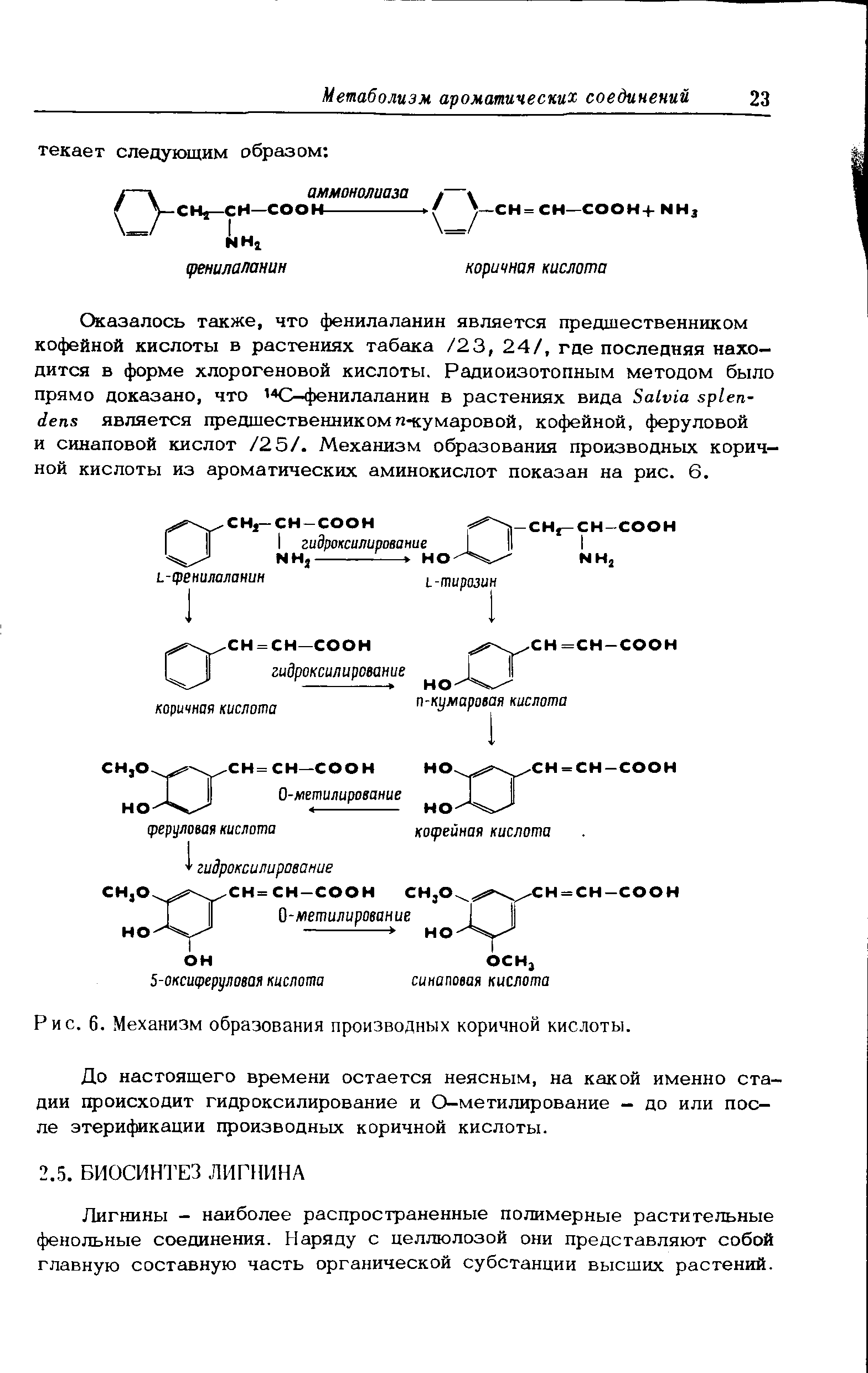
Оказалось также, что фенилаланин является предшественником кофейной кислоты в растениях табака /23, 24/, где последняя находится в форме хлорогеновой кислоты. Радиоизотопным методом было прямо доказано, что ,4С-фенилаланин в растениях вида Salvia splen-dens является предшественникомn-кумаровой, кофейной, феруловой и синаповой кислот /25/. Механизм образования производных коричной кислоты из ароматических аминокислот показан на рис. 6.~~Роль сахаров в процессе лигнификации выяснилась в последние годы. Исследованиями с меченой глюкозой древесины сосны Pinus strobus и Eucalyptus nitens /27, 28/ был подтвержден механизм образования лигнина через шикимовую (см. рис. 2) и префеновую (рис. 3) кислоты. Аналогичные эксперименты с меченой шикимовой кислотой подтвердили, что этот механизм является при лигнификации решающим. Ацетатный механизм (см. рис. 1) в процессе лигнификации не играет роли. Шикимовая кислота чрезвычайно распространена в высших растениях,и в настоящее время установлено, что шикимовая и префеновая кислоты управляют синтезом ароматических соединений в микроорганизмах и растениях. На рис. 5 показано, что первичными ароматическими соединениями, образующимися по этому механизму, служат фенилпропаноиды - фенилпировиноградная и п-окси-фенилпировиноградная кислоты. Далее было установлено, что ключевую роль при образовании лигнина играют аромаэтнеские аминокио-лоты - фенилаланин и тирозин, которые являются промежуточными продуктами метаболизма фенилпропаноидов (рис. 5). Опыты с мечеными фенилаланином и тирозином однозначно доказали, что все растения обладают способностью инкорпорировать фенилаланин в ароматической части лигнина. С другой стороны, тирозин не играет такой универсальной роли в процессе лигнификации растений. Поэтому предпочтительным оказывается механизм с участием фенилаланина, превращаемого дезаминированием в коричную кислоту (рис. 6), а не сходный механизм перехода тирозина в n-кумаровую кислоту.
Вернуться к оглавлению