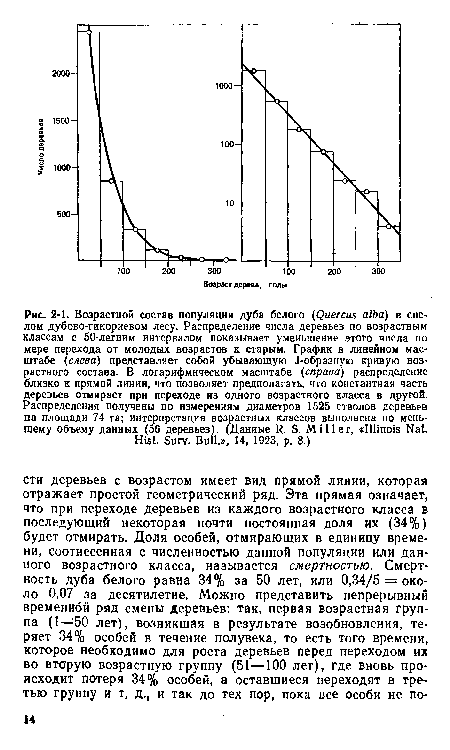
| Возрастной состав популяции дуба белого (Quercus alba) в спелом дубово-гикориевом лесу. Распределение числа деревьев по возрастным классам с 50-летним интервалом показывает уменьшение этого числа по мере перехода от молодых возрастов к старым. График в линейном масштабе {слева) представляет собой убывающую J-образную кривую возрастного состава. В логарифмическом масштабе (справа) распределение близко к прямой линии, что позволяет предполагать, что константная часть деревьев отмирает при переходе из одного возрастного класса в другой. Распределения получены по измерениям диаметров 1525 стволов деревьев на площади 74 га; интерпретация возрастных классов выполнена по меньшему объему данных (56 деревьев). (Данные R. S. Miller, «Illinois Nat. |
 |
Далее
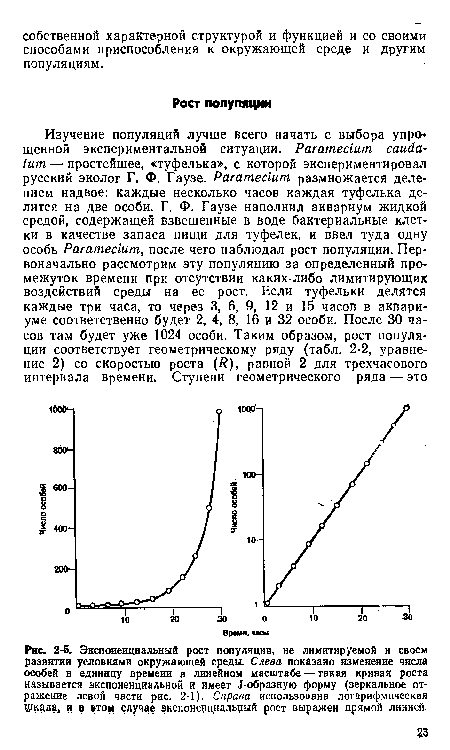
| Экспоненциальный рост популяции, не лимитируемой н своем развитии условиями окружающей среды. Слева показано изменение числа особей в единицу времени в линейном масштабе — такая кривая роста называется экспоненциальной и имеет Л-образную форму (зеркальное отражение левой части рис. 2-1). Справа использована логарифмическая щкдла, и в »том случае экспоненциальный рост выражен прямой линией. |
 |
Далее
| Ярусность и условия освещения в лесу. Различные виды деревьев, кустарников и трав, имеющие листья иа разной высоте над уровнем почвы (слева), адаптированы к неодинаковой интенсивности света (справа). Уменьшение освещенности является результатом поглощения солнечного света листвой. |
 |
Далее
| Четыре типа распределения популяции в сообществе. А. Случайное распределение (явно нерегулярное). В. Групповое, илн контагиозное. С. Равномерное, илн отрицательно контагиозное. £>. Комбинированное |
 |
Далее
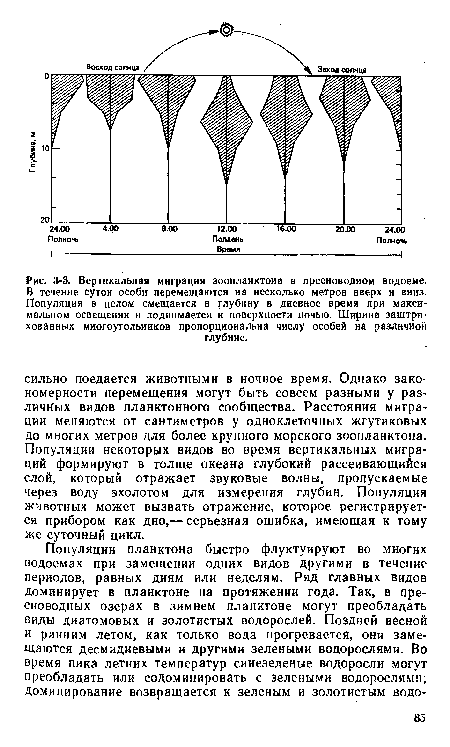
| Вер1икальная миграция зоопланктона в пресноводном водоеме. В течение суток особи перемещаются на несколько метров вверх и вниз. Популяция в целом смещается в глубину в дневное время при максимальном освещении и поднимается к поверхности ночью. Ширина заштрихованных многоугольников пропорциональна числу особей на различной |
 |
Далее
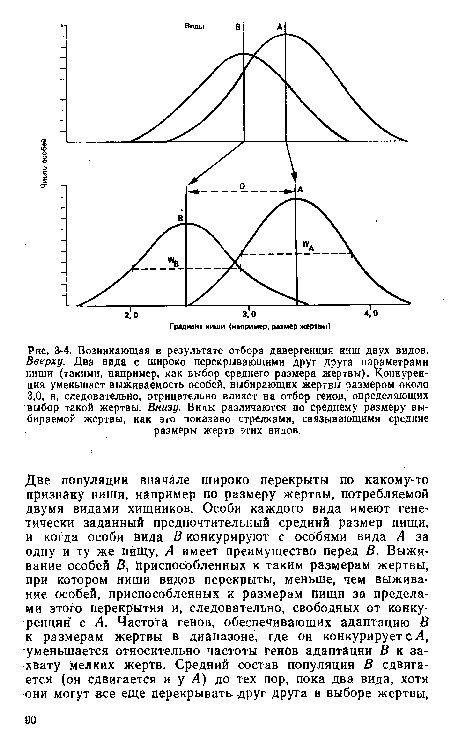
| Возникающая в результате отбора дивергенция ииш двух видов. Вверху. Два вида с широко перекрывающими друг друга параметрами ниши (такими, например, как выбор среднего размера жертвы). Конкуренция уменьшает выживаемость особей, выбирающих жертвы размером около 3,0, н, следовательно, отрицательно влияет на отбор геиов, определяющих выбор такой жертвы. Внизу. Виды различаются по среднему размеру выбираемой жертвы, как это показано стрелками, связывающими средние размеры жертв этих видов. |
 |
Далее
| Размещение ниш у восьми видов голубей, обитающих в тропическом дождевом лесу Новой Гвинеи и питающихся плодами. Четыре группы птиц показаны сидящими на деревьях, имеющих плодч четырех разных диаметров. Вес голубей в граммах проставлен над их изображениями; вес каждого голубя в пределах группы примерно в 1,5 раза больше веса соседнего с ним голубя этой же ииши. Каждое плодовое дерево привлекает четырех последовательных членов этого ряда. Деревья с более крупными плодами привлекают более крупных птиц. На данном дереве голуби меньшего размера питаются на ветках меньшего диаметра, то есть дальше от ствола. (J. М. Diamond. «Science», 179, 1973, p. 759—769.) |
 |
Далее
| Три гипотетические кривые значимости видов. Все кривые рассчитаны для гипотетической выборки, содержащей 20 видов. А. Гипотеза случайных границ между иишами. В. Геометрический ряд, с = 0,5. С. Логнормальное распределение (о способе изображения данных см. рис. 3-11) для природного сообщества. (R. Н. Whittaker. «Science», 147, 1965, |
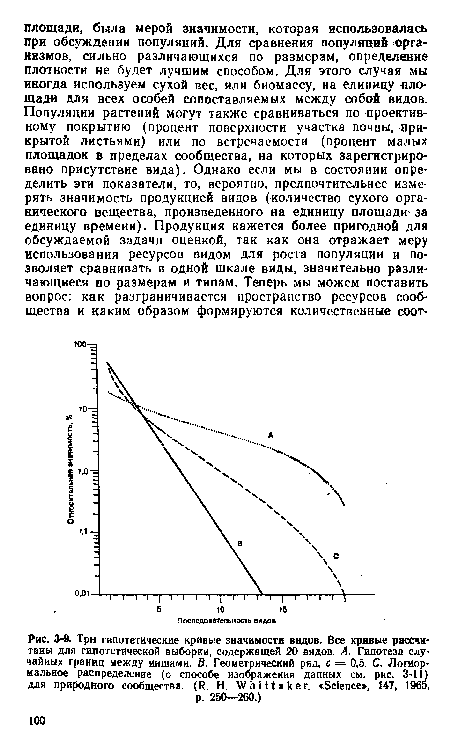 |
Далее
| Четыре гипотетических типа распределения видов по градиенту среды. Каждая кривая иа рисунке отражает распределение одной видовой |
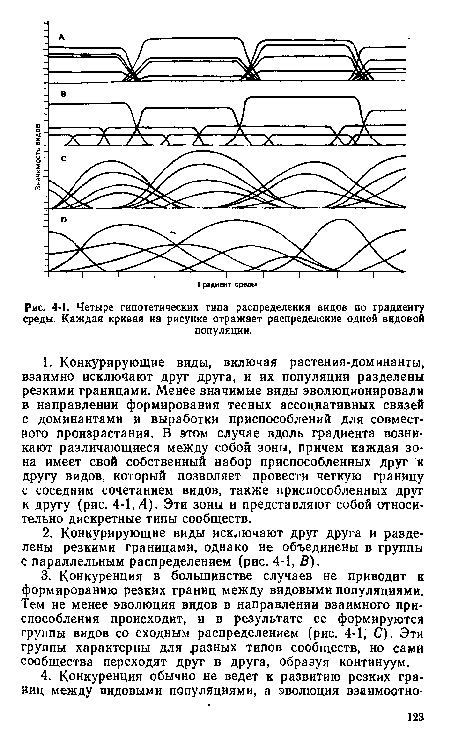 |
Далее
| Схема растительности гор Санта-Каталина (свыше 2800 м над уровнем моря), штат Аризона. Четыреста описаний растительности распределены на схеме в зависимости от их положения на осях градиента высоты над уровнем моря (слева) и градиента топографического увлажнения (внизу). Сплошные линии соединяют средние положения каждого типа сообществ. Доминаитные виды показаны для той части распределения, где они имеют наибольшую значимость. (R. Н. Whittaker, W. А. N i е г i n g, «Ecology», 46, 1965, p. 429—452.) |
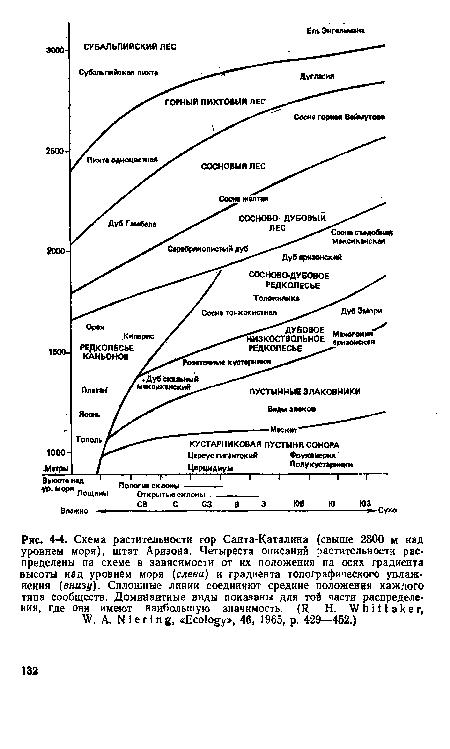 |
Далее
| Положение десяти лесных сообществ Польши в двух осях полярной ордииации. Каждое описание представляет собой среднее из нескольких полевых описаний, характерязукяцих тип сообщества (данные |
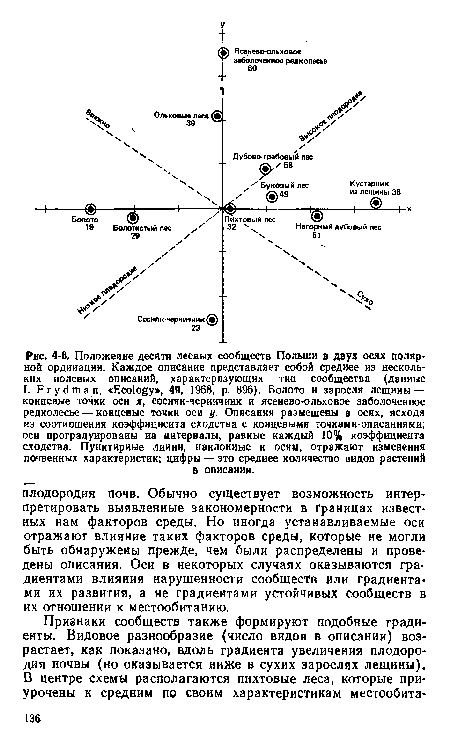 |
Далее
| Динамика популяций птиц в ходе сукцессии. Показаны виды, которые учтены в количестве не менее чем 15 пар на 100 акров территории (40 га) при восстановлении залежи в лес, район Пндмонта, штат Джорджия. |
 |
Далее
| Зависимость чистой первичной продукции надземных и подземных органов от среднегодовой температуры. (Н. L i е t h. «Human Ecol.», 1, |
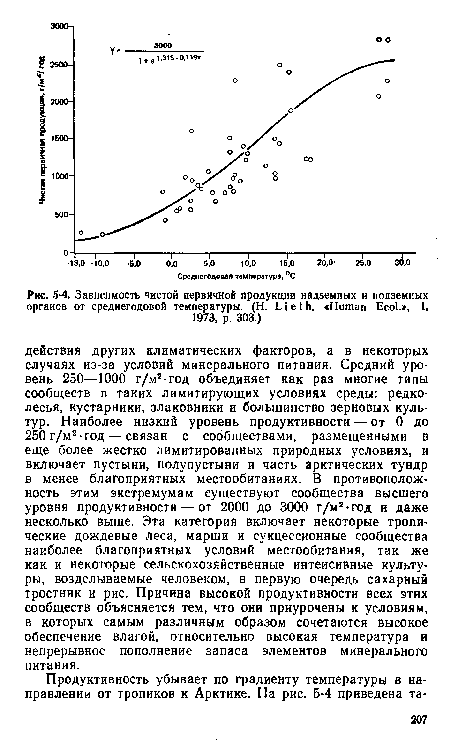 |
Далее
| Поток энергии в природном сообществе. Для устойчивого сообщества наблюдается баланс между поступлением в сообщество энергии фотосинтеза (слева) и ее рассеиванием в окружающую среду (справа). Фиксируемая в органическом веществе сообщества энергия остается постоянной. |
 |
Далее
| Профили трех типов почв. Слева |
 |
Далее