| Кривая число видов — площадь в растительной ассамблее подлеска в одном из лесов Северной Каролины [6]. |
![Кривая число видов — площадь в растительной ассамблее подлеска в одном из лесов Северной Каролины [6].](/static/pngsmall/820484928.png) |
Далее
| Широтный градиент разнообразия гнездящихся птиц [15]. |
![Широтный градиент разнообразия гнездящихся птиц [15].](/static/pngsmall/820484934.png) |
Далее
| Плотность видов ящериц и разных районах США [14]. |
|
Далее
| Видовое богатство птиц в характерных местообитаниях умеренной зоны (Tramer Е., Ecology, 1969, 50, 927—929). |
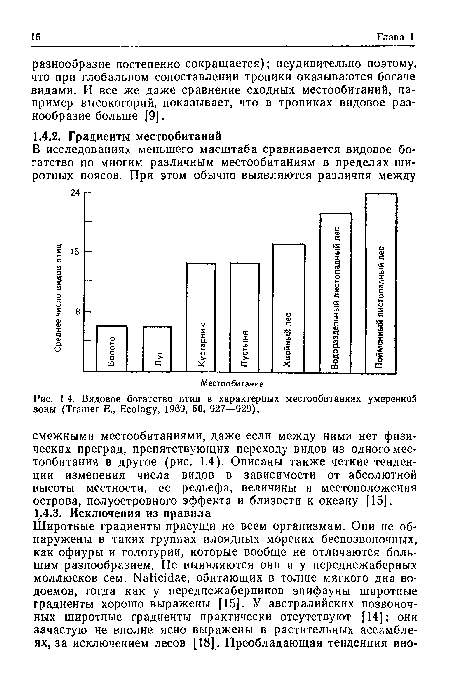 |
Далее
| Реакция вида на градиент одного из факторов среды. Приспособлен иость оценивается по успеху размножения, величине популяции и выживаемости. |
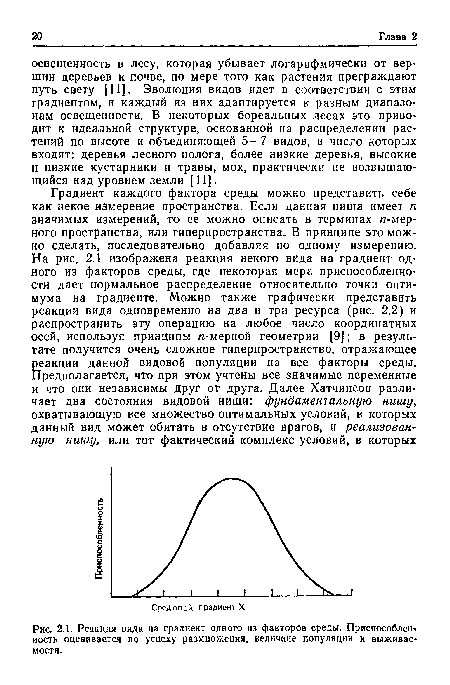 |
Далее
| Одновременная реакция вида на два (А) и иа три градиента факторов среды с учетом приспособленности вида (Б) [9]. |
![Одновременная реакция вида на два (А) и иа три градиента факторов среды с учетом приспособленности вида (Б) [9].](/static/pngsmall/820484946.png) |
Далее
| Возможные взаимоотношения между нишами двух видов вдоль одного градиента среды [9]. А. Включение одной ниши в другую. Б. Перекрывающиеся ниши. В. Соприкасающиеся ииши. Г. Разобщенные ниши. |
![Возможные взаимоотношения между нишами двух видов вдоль одного градиента среды [9]. А. Включение одной ниши в другую. Б. Перекрывающиеся ниши. В. Соприкасающиеся ииши. Г. Разобщенные ниши.](/static/pngsmall/820484952.png) |
Далее
| Сходство морфологических признаков у экологически равноценных видов птиц в Панаме (черные треугольники) и Либерии (белые треугольники) [43]. |
![Сходство морфологических признаков у экологически равноценных видов птиц в Панаме (черные треугольники) и Либерии (белые треугольники) [43].](/static/pngsmall/820484964.png) |
Далее
| Изменения ширины ниши в условиях конкуренции [46]. А Внутривидовая конкуренция — кривые потребления при низкой (/) и высокой (//) плотности популяции Б Межвидовая конкуренция Стрелки указывают направление изменений |
![Изменения ширины ниши в условиях конкуренции [46]. А Внутривидовая конкуренция — кривые потребления при низкой (/) и высокой (//) плотности популяции Б Межвидовая конкуренция Стрелки указывают направление изменений](/static/pngsmall/820484970.png) |
Далее
| Конкурентное исключение у мшанок. Стрелками показано направле ние нарастания [72], |
![Конкурентное исключение у мшанок. Стрелками показано направле ние нарастания [72],](/static/pngsmall/820484982.png) |
Далее
| Зависимость между средней величиной ниши и числом видов в выборках из зарослей пустынных кустарников (квадратики), злаковников (треугольнички) и смешанного леса (кружки) [6]. |
![Зависимость между средней величиной ниши и числом видов в выборках из зарослей пустынных кустарников (квадратики), злаковников (треугольнички) и смешанного леса (кружки) [6].](/static/pngsmall/820484990.png) |
Далее
| Теоретические кривые использования ресурсов для трех видов со сходными функциями использования (/ ), й — расстояние между средними значениями; и> — стандартное отклонение. |
 |
Далее
| Механизмы разделения ниш у лесных видов птиц [131]. А. Пищевая специализация без пространственного разделения. Б. Разделение по вертикали. В. Разделение по горизонтали. |
![Механизмы разделения ниш у лесных видов птиц [131]. А. Пищевая специализация без пространственного разделения. Б. Разделение по вертикали. В. Разделение по горизонтали.](/static/pngsmall/820485026.png) |
Далее
| Распределение ресурсов между |
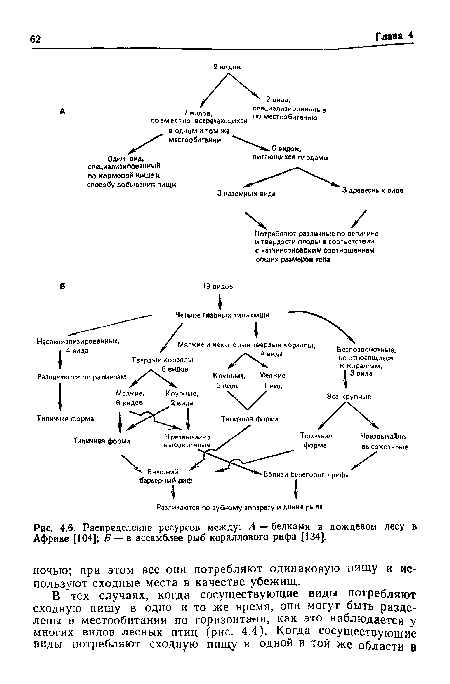 |
Далее
| Сукцессионные взаимоотношения между типами растительности на хорошо дренируемых кислых почвах водоразделов Великобритании при низкой или незначительной (А) и при сильной пастбищной нагрузке (Б). Жирными стрелками показаны обычные последовательности, а тонкими — более редкие [4]. |
![Сукцессионные взаимоотношения между типами растительности на хорошо дренируемых кислых почвах водоразделов Великобритании при низкой или незначительной (А) и при сильной пастбищной нагрузке (Б). Жирными стрелками показаны обычные последовательности, а тонкими — более редкие [4].](/static/pngsmall/820485054.png) |
Далее
| Трофические взаимосвязи в пищевой подсети залива Макко-Бей (шт. Вашингтон) Жирной стрелкой показано сильное взаимодействие [190]. |
![Трофические взаимосвязи в пищевой подсети залива Макко-Бей (шт. Вашингтон) Жирной стрелкой показано сильное взаимодействие [190].](/static/pngsmall/820485064.png) |
Далее
| Изменения разнообразия и сложности (низкая или высокая степень) при различном числе взаимодействующих уровней. Стрелками показано направление взаимодействий; сплошными стрелками — сильные взаимодействия, а пунктирной — слабые. Сидячие беспозвоночные и бентосные вододосли отнесены к одному и тому же уровню, так как они конкурируют за пространство. Цифры в скобках — ссылки на литературу. |
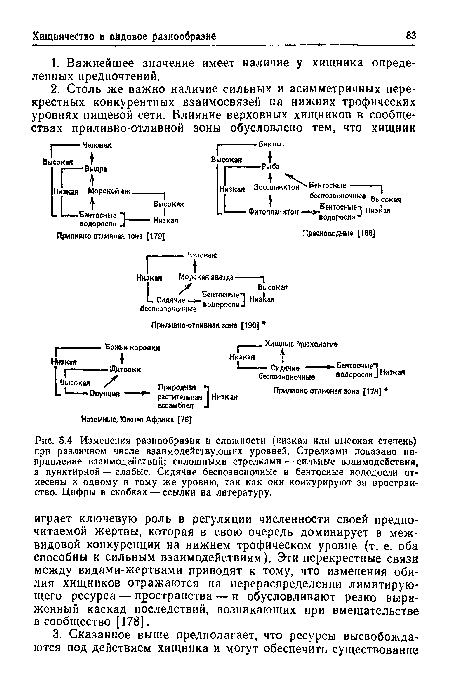 |
Далее
| Предложенная Коннеллом [195] классификация механизмов организации сообщества, основанная на воздействиях внешних факторов. А. Популяции, численность которых ограничивается неблагоприятными физическими факторами. Б. Популяции, численность которых ограничивается интенсивным хищничеством. |
![Предложенная Коннеллом [195] классификация механизмов организации сообщества, основанная на воздействиях внешних факторов. А. Популяции, численность которых ограничивается неблагоприятными физическими факторами. Б. Популяции, численность которых ограничивается интенсивным хищничеством.](/static/pngsmall/820485080.png) |
Далее
| Модели процесса колонизации островов материковыми видами. А. Равновесная модель Мак-Артура и Уилсона [28]. Б. Модель постоянных обитателей [203]. В. Комбинация моделей Л и />. [203]. Г. Модель, предсказывающая число видов (Б) и среднюю величину популяций (Ы) для ящериц и птиц иа мелких островах [204]. Д. Стадии колонизации островов материковыми видами [205]. 5 — равновесное число видов; Р — величина видового фонда; / — кривая иммиграции // — кривая вымирания. |
![Модели процесса колонизации островов материковыми видами. А. Равновесная модель Мак-Артура и Уилсона [28]. Б. Модель постоянных обитателей [203]. В. Комбинация моделей Л и />. [203]. Г. Модель, предсказывающая число видов (Б) и среднюю величину популяций (Ы) для ящериц и птиц иа мелких островах [204]. Д. Стадии колонизации островов материковыми видами [205]. 5 — равновесное число видов; Р — величина видового фонда; / — кривая иммиграции // — кривая вымирания.](/static/pngsmall/820485094.png) |
Далее
| Зависимости между числом видов и площадью «острова», представленного растением-хозяином. А Насекомые, паразитирующие на деревьях (/), кустарниках (//) и травянистых растениях (III) Британских островов [217]. 6 Пилильщики-минеры и деревья Британских островов [218]. |
![Зависимости между числом видов и площадью «острова», представленного растением-хозяином. А Насекомые, паразитирующие на деревьях (/), кустарниках (//) и травянистых растениях (III) Британских островов [217]. 6 Пилильщики-минеры и деревья Британских островов [218].](/static/pngsmall/820485106.png) |
Далее
| Зависимости число видов — площадь на материках А. Виды цветковых растений в обследованных районах Англии [61] Б. Птицы Северной Америки [293]. |
![Зависимости число видов — площадь на материках А. Виды цветковых растений в обследованных районах Англии [61] Б. Птицы Северной Америки [293].](/static/pngsmall/820485110.png) |
Далее
| Две модели насыщения видами материков. А. Модель Мак-Артура [28]. Б. Альтернативная модель, основанная на взглядах Майра [225]. I — кривая видообразования; II — кривая вымирания; 5 — равновесное число видов. |
![Две модели насыщения видами материков. А. Модель Мак-Артура [28]. Б. Альтернативная модель, основанная на взглядах Майра [225]. I — кривая видообразования; II — кривая вымирания; 5 — равновесное число видов.](/static/pngsmall/820485112.png) |
Далее
| Примеры насыщения видами. Л. Сравнимость а-разнообразии ассамблей птиц в трех областях средиземноморского типа [42]. Б. Сравнимость зависимости между разнообразием видов птнц и разнообразием деревьев по высоте расположения листвы в Австралии (квадратики) и в Северной Америке (треугольнички) [28]. |
![Примеры насыщения видами. Л. Сравнимость а-разнообразии ассамблей птиц в трех областях средиземноморского типа [42]. Б. Сравнимость зависимости между разнообразием видов птнц и разнообразием деревьев по высоте расположения листвы в Австралии (квадратики) и в Северной Америке (треугольнички) [28].](/static/pngsmall/820485114.png) |
Далее
| Включение новых видов в сообщество (Л) происходит при помощи нескольких механизмов |
 |
Далее
| Схема гипотезы стабильность — время Сандерса [230]. |
![Схема гипотезы стабильность — время Сандерса [230].](/static/pngsmall/820485132.png) |
Далее
| Зависимости между колебаниями температуры и видовым богатством птиц (А) и брюхоногих моллюсков (Б) западного побережья Америки [299] |
|
Далее
| Гипотеза промежуточного нарушения [137]. |
![Гипотеза промежуточного нарушения [137].](/static/pngsmall/820485136.png) |
Далее
| Зависимость между числом грызунов и продуктивностью на песчаных равнинах в пустыне Сонора (I и квадраты) и на песчаных дюнах в пустыне Мохаве и Большом Бассейне (II и кружки) [58]. |
![Зависимость между числом грызунов и продуктивностью на песчаных равнинах в пустыне Сонора (I и квадраты) и на песчаных дюнах в пустыне Мохаве и Большом Бассейне (II и кружки) [58].](/static/pngsmall/820485146.png) |
Далее
| Гипотеза кольцевой сети. А. Обычная линейная конкурентная иерархия. Б. Кольцевая конкурентная иерархия. В, Ослабление. |
 |
Далее
| Распределение малайских бабочек по частоте [253]. |
![Распределение малайских бабочек по частоте [253].](/static/pngsmall/820485164.png) |
Далее
| Примеры логарифмически нормального распределения обилия видов, обнаруженного на основании |
 |
Далее
| Каноническое логарифмически нормальное распределение для ассамблей из 178 видов птиц, построенное таким образом, чтобы средняя точка совпадала с нулем (1 — виды; II — особи [255]). |
![Каноническое логарифмически нормальное распределение для ассамблей из 178 видов птиц, построенное таким образом, чтобы средняя точка совпадала с нулем (1 — виды; II — особи [255]).](/static/pngsmall/820485168.png) |
Далее
| Две возможные последовательности разбиений в модели минимальной структуры сообщества, предложенной Сугихарой, и соответствующие данные для ассамблей реальных видов [2101. |
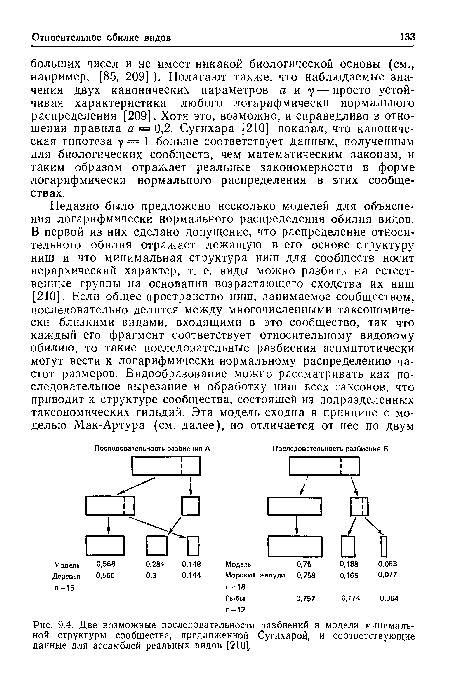 |
Далее
| Примеры распределения видового обилия по принципу «разломанной палки». А. Ассамблея рыб (сем. РегЫс1ае). Б. Офиуры; прерывистая линия — ожидаемое, сплошная — наблюдаемое распределение [259]. |
![Примеры распределения видового обилия по принципу «разломанной палки». А. Ассамблея рыб (сем. РегЫс1ае). Б. Офиуры; прерывистая линия — ожидаемое, сплошная — наблюдаемое распределение [259].](/static/pngsmall/820485174.png) |
Далее
| Графики распределения обилия видов, предсказываемого четырьмя различными моделями |
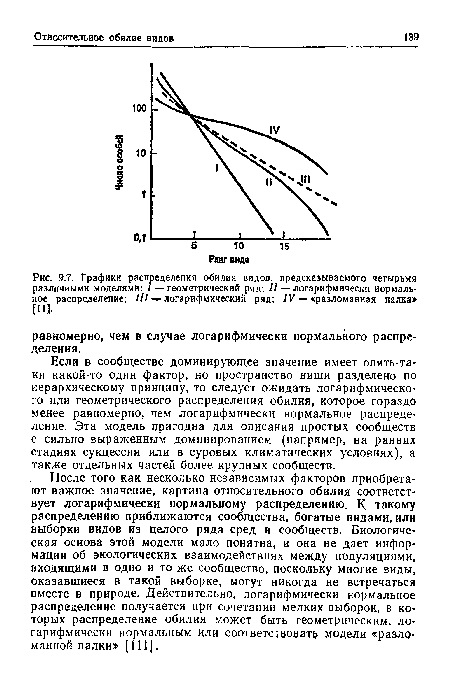 |
Далее
| Распределение обилия видов в процессе сукцессии от залежи к сложному бору. Для исходного залежного сообщества (/) характерно геометрическое распределение. На более поздней стадии травяной растительности (2) распределение приближается к логарифмически нормальному На стадии (3) в ассамблею проникают древесные растения, которые постепенно становятся мощными доминантами (4), что ведет к понижению видового разнообразия. Кривая (5) отражает начало вторичного возрастания видового разнообразия [И]. |
![Распределение обилия видов в процессе сукцессии от залежи к сложному бору. Для исходного залежного сообщества (/) характерно геометрическое распределение. На более поздней стадии травяной растительности (2) распределение приближается к логарифмически нормальному На стадии (3) в ассамблею проникают древесные растения, которые постепенно становятся мощными доминантами (4), что ведет к понижению видового разнообразия. Кривая (5) отражает начало вторичного возрастания видового разнообразия [И].](/static/pngsmall/820485184.png) |
Далее
| Среднее разделение ниш по трем измерениям в десяти ассамблеях луговых птиц Квадраты — южноамериканские ассамблеи, кружки — североамериканские [135]. |
![Среднее разделение ниш по трем измерениям в десяти ассамблеях луговых птиц Квадраты — южноамериканские ассамблеи, кружки — североамериканские [135].](/static/pngsmall/820485192.png) |
Далее
| Зависимость между плотностью видов ящериц и общим разнообразием ресурсов, используемых по трем измерениям ниши (пища, микроэкотоп и время активности). Квадратики — Северная Америка, треугольники — пустыня Калахари (Африка), кружки — Австралия [41]. |
![Зависимость между плотностью видов ящериц и общим разнообразием ресурсов, используемых по трем измерениям ниши (пища, микроэкотоп и время активности). Квадратики — Северная Америка, треугольники — пустыня Калахари (Африка), кружки — Австралия [41].](/static/pngsmall/820485202.png) |
Далее
| Зависимость между логарифмом числа видов хищников и паразито-идов и логарифмом числа потенциальных видов-жертв на деревьях в Южной Африке (белые кружки) и в Великобритании (черные кружки) [250]. |
![Зависимость между логарифмом числа видов хищников и паразито-идов и логарифмом числа потенциальных видов-жертв на деревьях в Южной Африке (белые кружки) и в Великобритании (черные кружки) [250].](/static/pngsmall/820485214.png) |
Далее