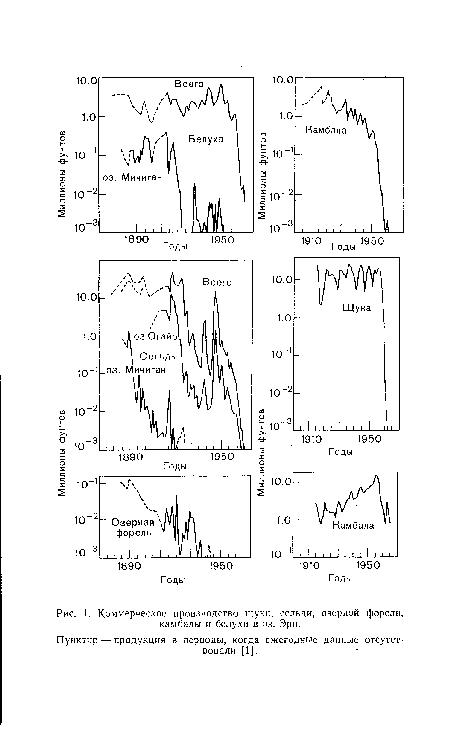
| Коммерческое производство щуки, сельди, озерной форели, камбалы и белухи в оз. Эри. |
 |
Далее
| Проектное развитие источников электроэнергии в США [4]. |
![Проектное развитие источников электроэнергии в США [4].](/static/pngsmall/389824932.png) |
Далее
| За. Экспоненциальный рост населения, потребление энергии и производство удобрений [3]. |
![За. Экспоненциальный рост населения, потребление энергии и производство удобрений [3].](/static/pngsmall/389824940.png) |
Далее
| Применение удобрений (Ы+РгСЬ+КгО) на единицу поверхности пахотной земли в зависимости от плотности населения [3]. |
![Применение удобрений (Ы+РгСЬ+КгО) на единицу поверхности пахотной земли в зависимости от плотности населения [3].](/static/pngsmall/389824942.png) |
Далее
| Равновесие между фотосинтезом (Ф) и дыханием (Д). Нарушение равновесия Ф—Д является следствием вертикального (озеро) или продольного (река) разделения Ф и Д организмов. Нарушение равновесия между фотосинтезом и дыханием ведет к загрязняющему влиянию того или иного вида |
 |
Далее
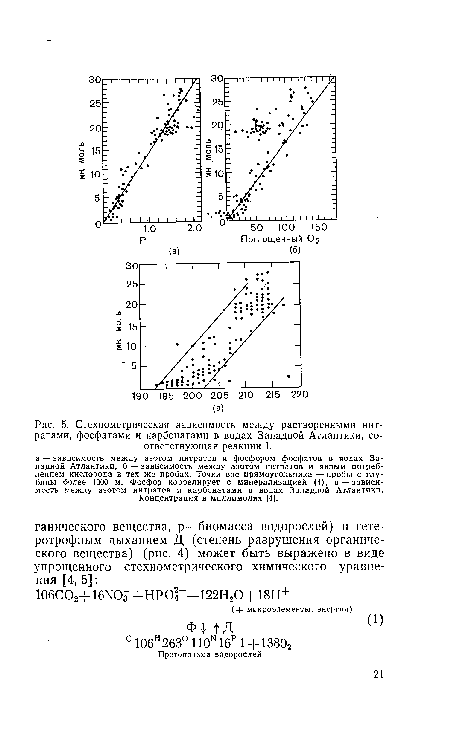
| Стехиометрическая зависимость между растворенными нитратами, фосфатами и карбонатами в водах Западной Атлантики, соответствующая реакции I. |
 |
Далее
| Изучение профиля фосфатов в бассейне Чарлз-Ривер. |
 |
Далее
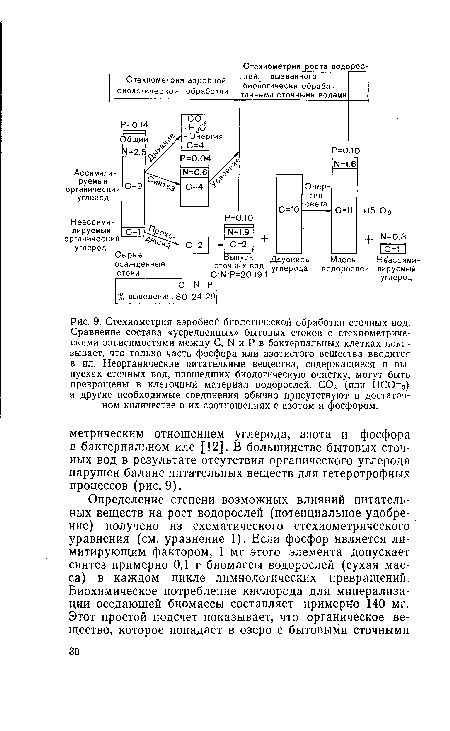
| Стехиометрия аэробной биологической обработки сточных вод. Сравнение состава «усредненных» бытовых стоков с стехиометриче-скими зависимостями между С, N и Р в бактериальных клетках показывает, что только часть фосфора или азотистого вещества вводится в ил. Неорганические питательные вещества, содержащиеся в выпусках сточных вод, прошедших биологическую очистку, могут быть превращены в клеточный материал водорослей. С02 (или НСО |
 |
Далее
| Превращения фосфата [14]. Схематически изображены локализация фосфора в воде и обменные процессы между различными локализациями. Hayes и Phillips [22] предположили, что время цикла (время, необходимое для потери из фазы всего имеющегося фосфора) колеблется от 5 мин для обмена между растворенным неорганическим фосфатом и фитопланктоном до многих дней для некоторых других обменов. Самые медленные процессы, по-видимому,— это обмен между водой и осадком ( |
![Превращения фосфата [14]. Схематически изображены локализация фосфора в воде и обменные процессы между различными локализациями. Hayes и Phillips [22] предположили, что время цикла (время, необходимое для потери из фазы всего имеющегося фосфора) колеблется от 5 мин для обмена между растворенным неорганическим фосфатом и фитопланктоном до многих дней для некоторых других обменов. Самые медленные процессы, по-видимому,— это обмен между водой и осадком (](/static/pngsmall/389824968.png) |
Далее
| Упрощенная модель равновесного состояния, характеризующего главные ступени лимнологического превращения фосфата в озере [18]. Модель воспроизводит реальную систему с-гипотетическим равновесием избытка различных форм фосфора (числа в прямоугольниках обозначают микрограммы Р на 1 л озерной воды) и скоростей обмена (числа под стрелками обозначают микрограммы Р на 1 л озерной воды в день). Цикл фосфора зависит главным образом от восстановления Р из биоты. Первичная продукция зависит от запаса Р в трофическом слое. Для более глубоких озер скорость поставки из осадков мала по сравнению с поставкой гиполим-ниона и введением Р из стоков и дренажа. Значительная часть Р, введенного в озеро, безвозвратно теряется в осадках. |
![Упрощенная модель равновесного состояния, характеризующего главные ступени лимнологического превращения фосфата в озере [18]. Модель воспроизводит реальную систему с-гипотетическим равновесием избытка различных форм фосфора (числа в прямоугольниках обозначают микрограммы Р на 1 л озерной воды) и скоростей обмена (числа под стрелками обозначают микрограммы Р на 1 л озерной воды в день). Цикл фосфора зависит главным образом от восстановления Р из биоты. Первичная продукция зависит от запаса Р в трофическом слое. Для более глубоких озер скорость поставки из осадков мала по сравнению с поставкой гиполим-ниона и введением Р из стоков и дренажа. Значительная часть Р, введенного в озеро, безвозвратно теряется в осадках.](/static/pngsmall/389824970.png) |
Далее
| Критическая нагрузка фосфором как функция глубины [3]. |
![Критическая нагрузка фосфором как функция глубины [3].](/static/pngsmall/389824974.png) |
Далее
| Направления в экологической преемственности. Экологические системы имеют тенденцию к переходу в стационарное состояние и постепенному самовосстановлению относительно энергии и веществ. Большинство характерных признаков приведено по Odum [1]. |
![Направления в экологической преемственности. Экологические системы имеют тенденцию к переходу в стационарное состояние и постепенному самовосстановлению относительно энергии и веществ. Большинство характерных признаков приведено по Odum [1].](/static/pngsmall/389824984.png) |
Далее
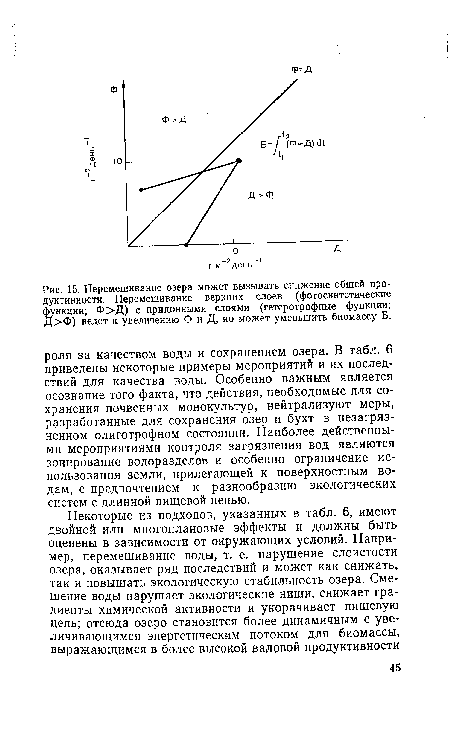
| Перемешивание озера может вызывать снижение общей продуктивности. Перемешивание верхних слоев (фотосинтетические функции; Ф>Д) с придонными слоями (гетеротрофные функции; Д>Ф) ведет к увеличению Ф и Д, но может уменьшить биомассу Б. |
 |
Далее
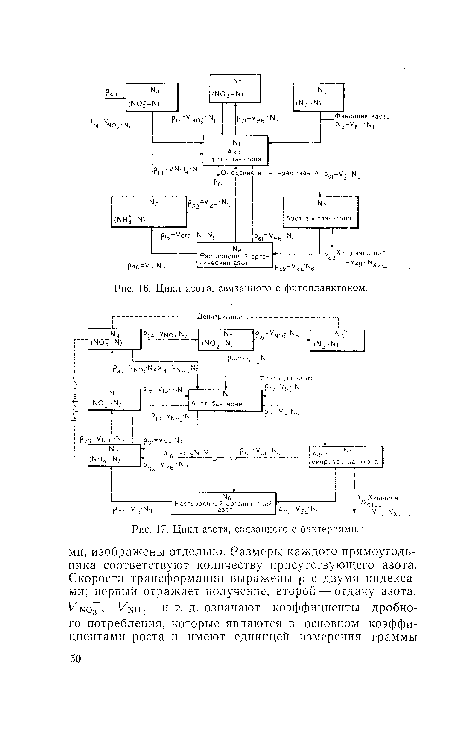
| Цикл азота, связанного с бактериями. |
 |
Далее
| Упрощенная схема превращения основного питательного вещества в эвтрофической зоне (пути регенерации опущены). |
 |
Далее
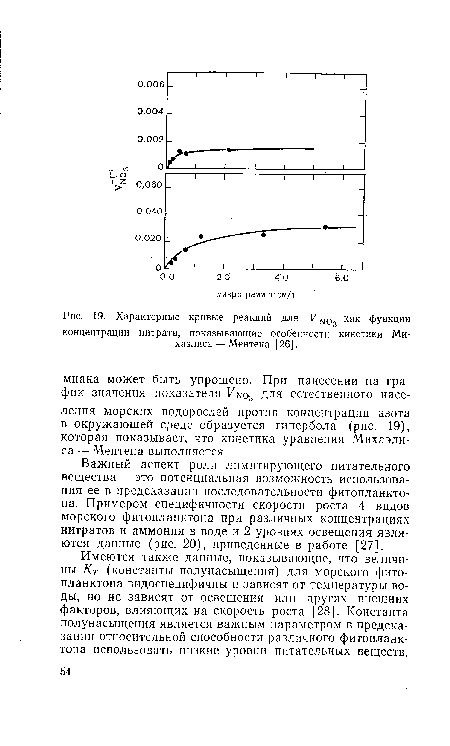
| Характерные кривые реакций для функции |
 |
Далее
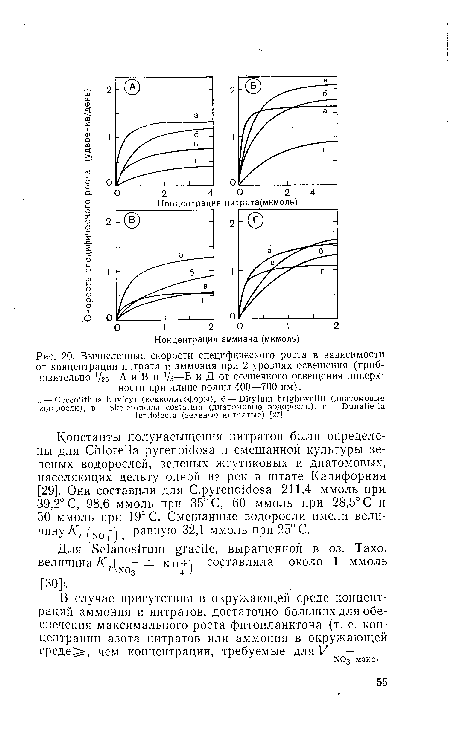
| Вычисленные скорости специфического роста в зависимости от концентрации нитрата и аммония при 2 уровнях освещения (приблизительно V20-—А и В и J/5—Б и Д от солнечного освещения поверх ности при длине волны 400—700 нм). |
 |
Далее
| Схематичное изображение возможного пути окисления железа Т. {еггоох1с1ап5 (модифицировано по [34]). |
![Схематичное изображение возможного пути окисления железа Т. {еггоох1с1ап5 (модифицировано по [34]).](/static/pngsmall/389825058.png) |
Далее
| Свободная энергия, освобожденная на электрон-моль для различных органических доноров электронов. Реактаиты и продукты реакции при единице активности, кроме [Н+] = [ОН |
![Свободная энергия, освобожденная на электрон-моль для различных органических доноров электронов. Реактаиты и продукты реакции при единице активности, кроме [Н+] = [ОН](/static/pngsmall/389825094.png) |
Далее
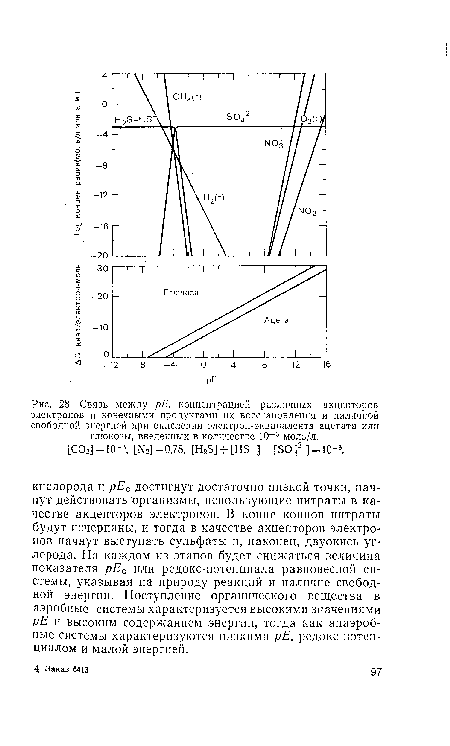
| Связь между рЕ, концентрацией различных акцепторов электронов и конечными продуктами их восстановления и наличной свободной энергией при окислении электрон-эквивалента ацетата или глюкозы, введенных в количестве 10 |
 |
Далее
| Равновесие концентрации различных компонентов (в электрон-эквивалентах) в зависимости от степени разведения в процессе анаэробного метанового брожения или сточных вод при 35°С [18]. |
![Равновесие концентрации различных компонентов (в электрон-эквивалентах) в зависимости от степени разведения в процессе анаэробного метанового брожения или сточных вод при 35°С [18].](/static/pngsmall/389825120.png) |
Далее
| Специфическая скорость удаления органических загрязнителей двумя типами фитоценозов как функция скорости течения. Эксперименты на модельных реках, питаемых подземной водой при одинаковых гидравлических условиях. Загрязнение |
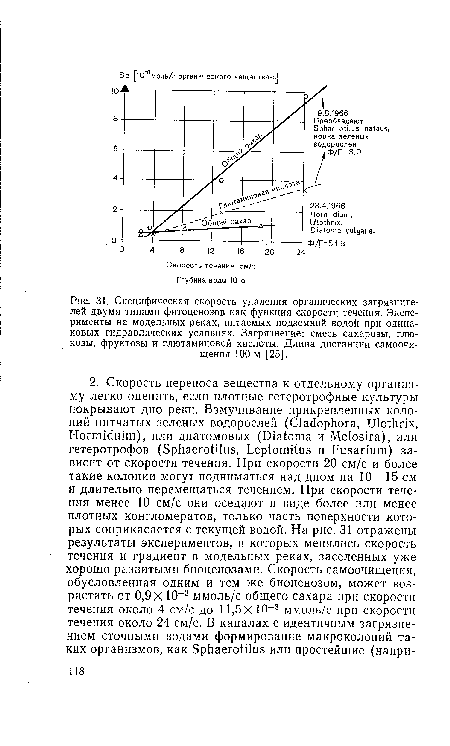 |
Далее
| Кривые скорости отмирания колиформных организмов в различных реках [14, 16]. |
![Кривые скорости отмирания колиформных организмов в различных реках [14, 16].](/static/pngsmall/389825150.png) |
Далее
| Схемы продукции и потребления биомассы вдоль профиля загрязненной реки. Расположение А=Е, как на рис. 37. |
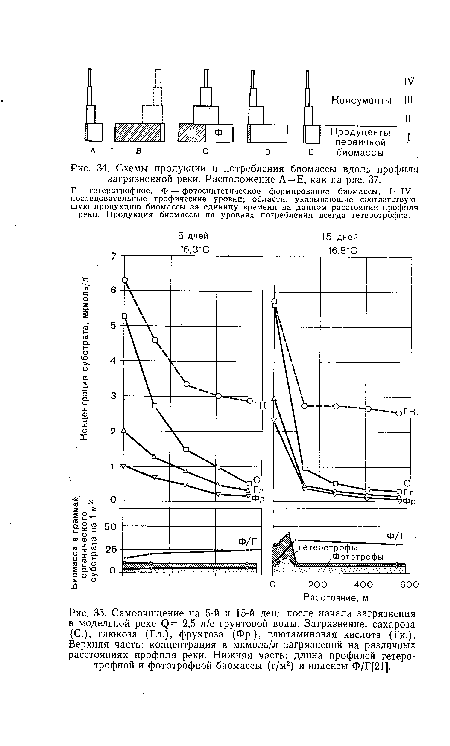 |
Далее
| Самоочищение на 5-й и 15-й день после начала загрязнения в модельной реке (¡) = 2,5 л/с грунтовой воды. Загрязнение |
|
Далее
| Скорость специфического выведения сахарозы фитоценозом с различными индексами Ф/Г как функция концентрации сахарозы в проточной воде. По данным экспериментов, приведенным на рис. |
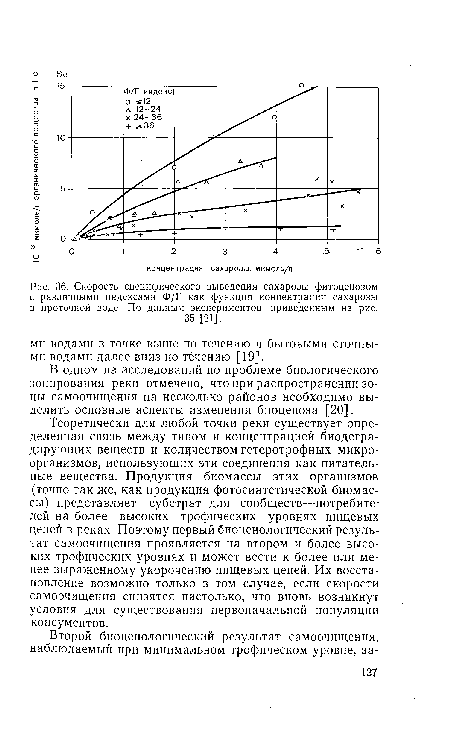 |
Далее
| Индекс Ф/Г фитоценоза в модельных реках при аналогичных |
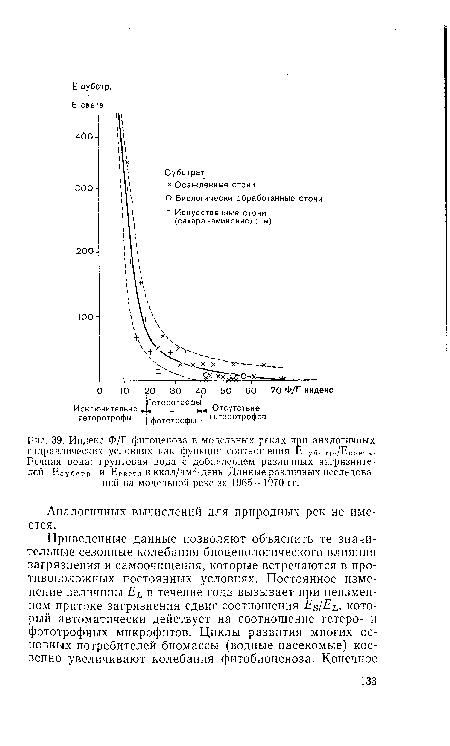 |
Далее
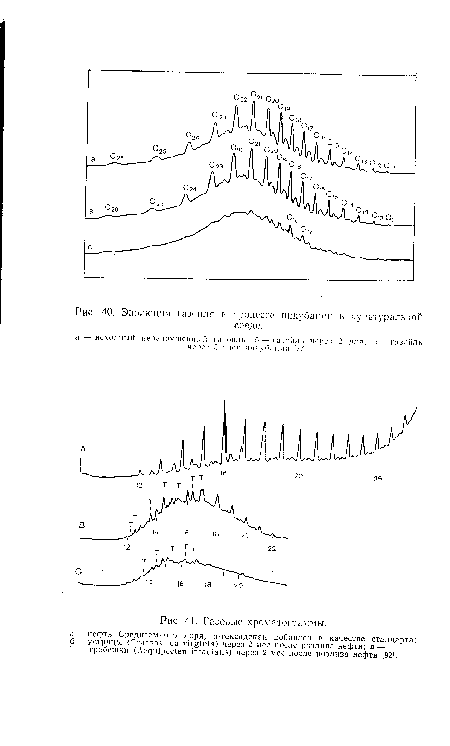
| Газовые хроматограммы. |
 |
Далее
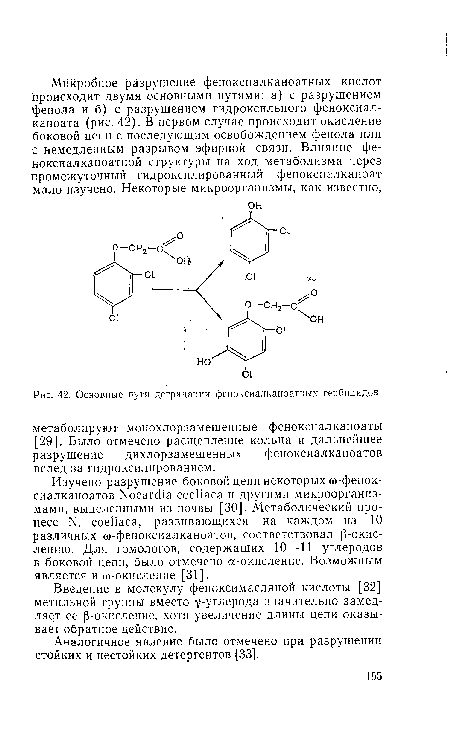
| Основные пути деградации феноксиалканоатных гербицидов. |
 |
Далее
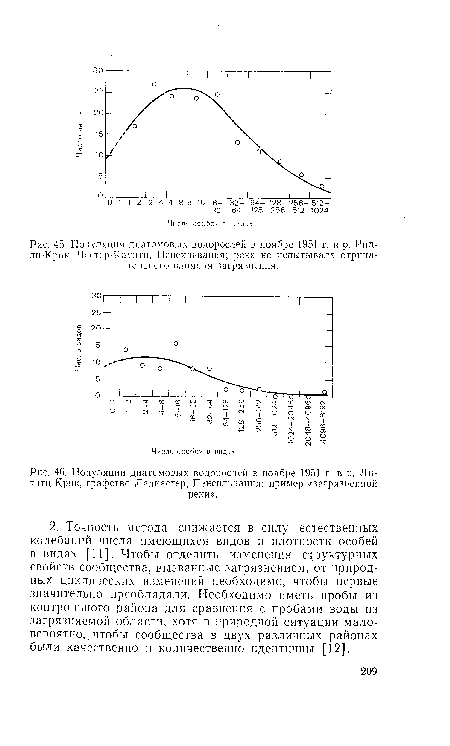
| Популяция диатомовых водорослей в ноябре 1951 г. в р. Ридли-Крик, Чсстчр-Кауити, Пенсильвания; река не испытывала отрицательного влияния загрязнения. |
 |
Далее
| Популяции диатомовых водорослей в ноябре 1951 г. в р. Ли-тнтц-Крик, графство Ланкастер, Пенсильвания; пример «загрязненной |
|
Далее
| Структура популяции водных сообществ при различных степенях загрязнения. Столбец I |
 |
Далее
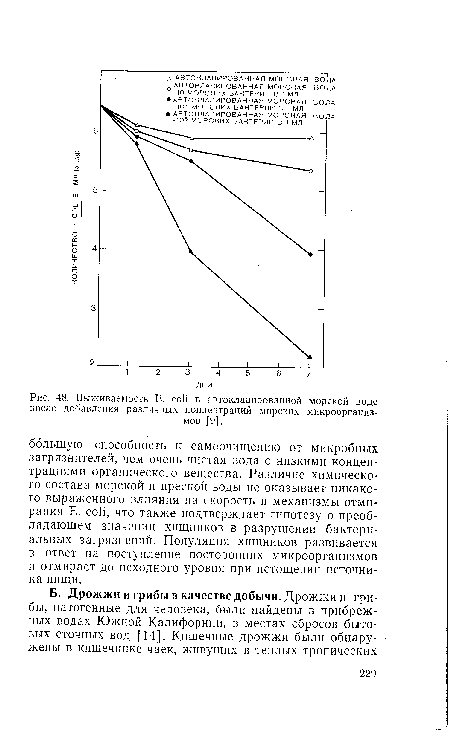
| Выживаемость E. coli в автоклавированной морской воде |
 |
Далее
| Взаимосвязь между количеством бактерий в речной воде и выживаемостью в ней бактериофага фх=174 [10]. |
![Взаимосвязь между количеством бактерий в речной воде и выживаемостью в ней бактериофага фх=174 [10].](/static/pngsmall/389825374.png) |
Далее
| Взаимосвязь между потреблением 02, удалением органического углерода и микробным ростом. |
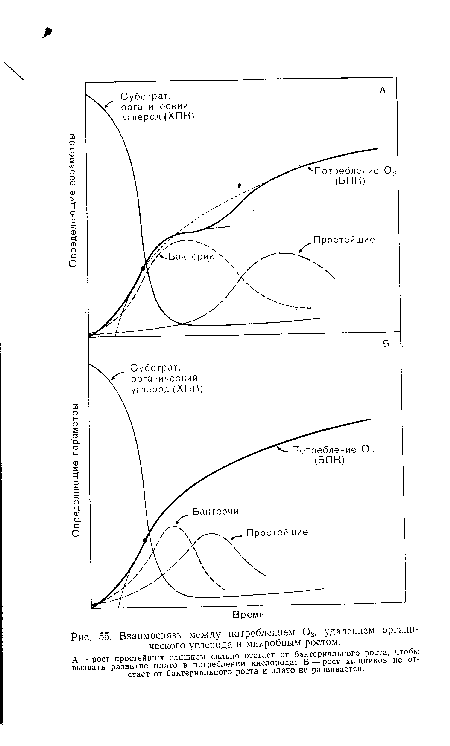 |
Далее
| Стандарты содержания органического вещества в выпусках очистных сооружений и в питьевой воде по сравнению с лимитирующими рост концентрациями глюкозы для E. coli в аэробных условиях |
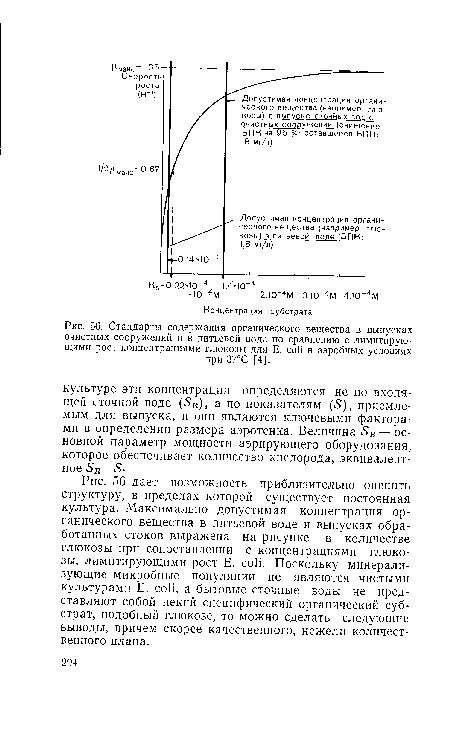 |
Далее
| Основные категории процессов микробиологической обработки органических стоков. |
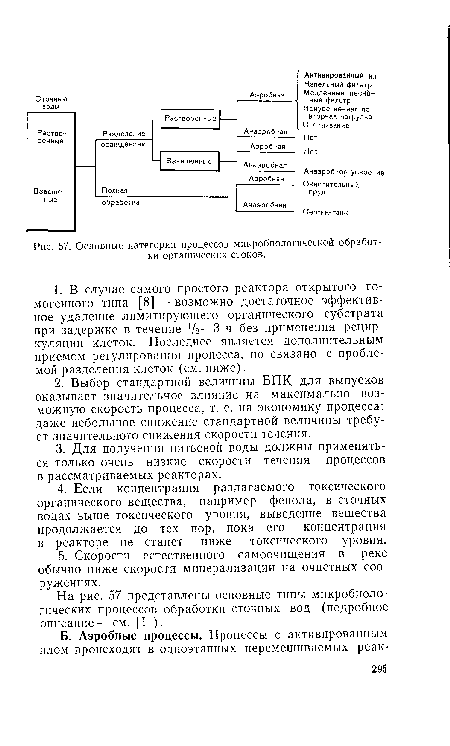 |
Далее
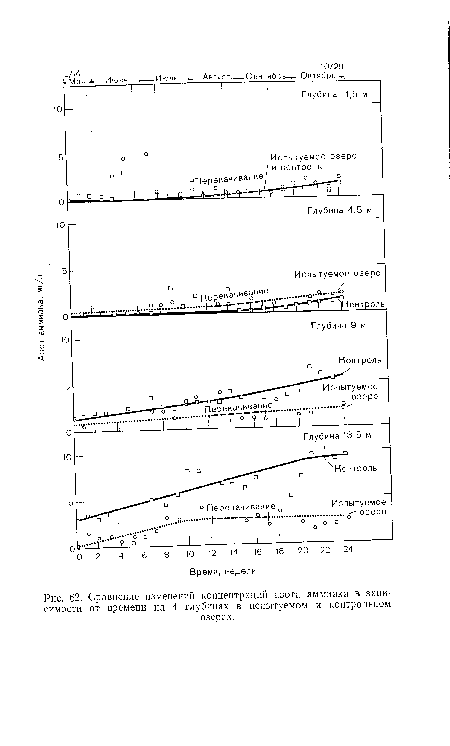
| Сравнение изменений концентрации растворенного кислорода в зависимости от времени на 4 глубинах в испытуемом и контрольном |
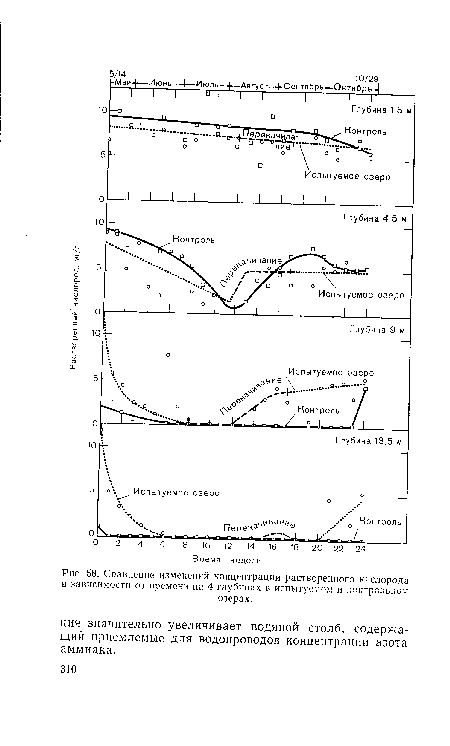 |
Далее
| Влияние воздухраспыляющего насоса на концентрацию растворенного кислорода. |
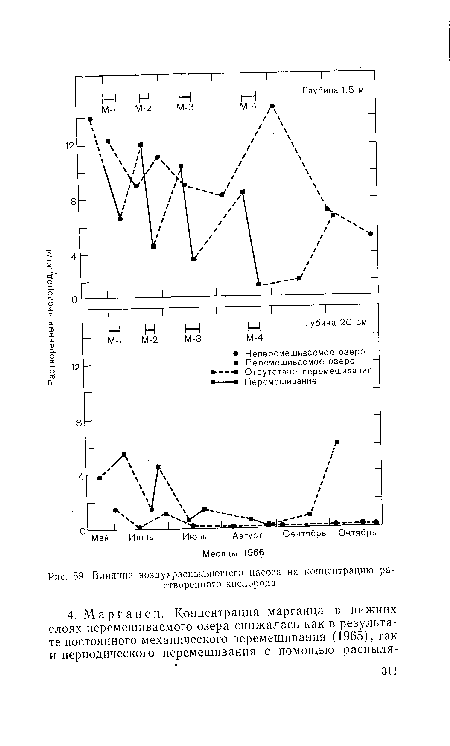 |
Далее
| Сравнение изменений концентраций сульфида в зависимости от времени на глубине 14 м в испытуемом и контрольном озерах. |
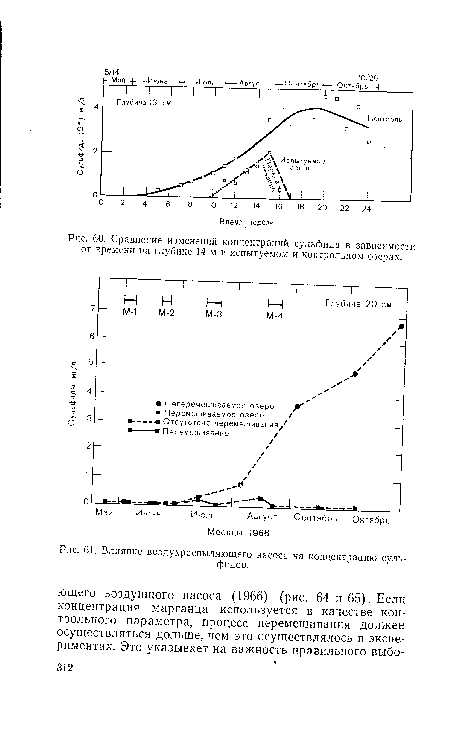 |
Далее
| Влияние воздухраспыляющего насоса на концентрацию сульфидов. |
|
Далее
| Сравнение изменений концентраций азота аммиака в зависимости от времени па 4 глубинах в испытуемом и контрольном |
 |
Далее
| Влияние воздухраспыляющего насоса на концентрацию |
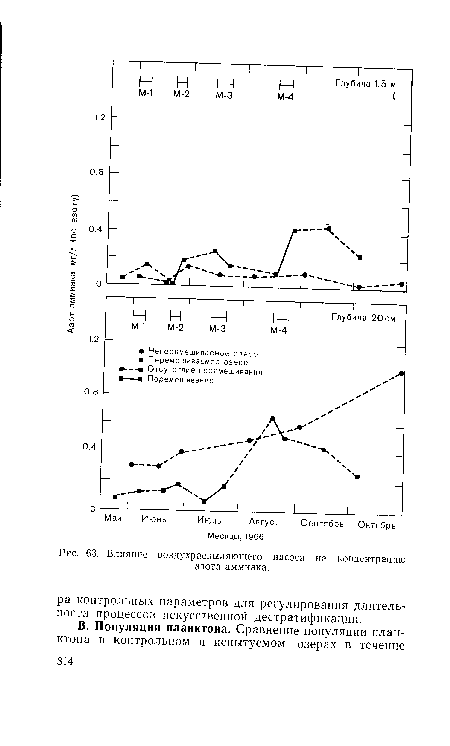 |
Далее
| Сравнение изменений общей концентрации марганца в зависимости от времени па двух глубинах в испытуемом и контрольном |
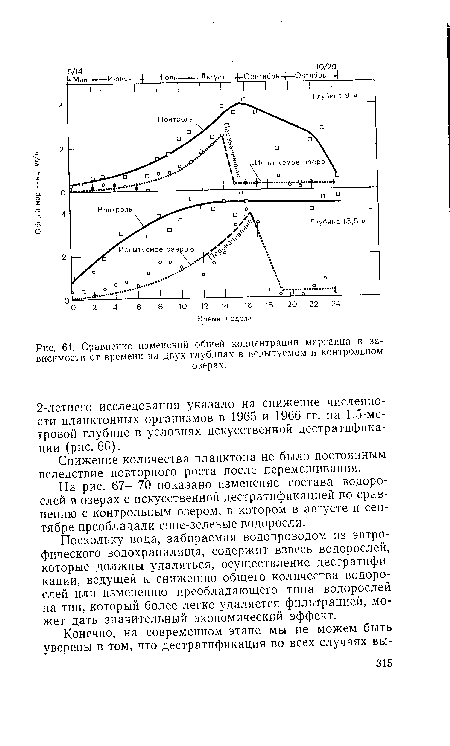 |
Далее
| Влияние воздухраспыляющего насоса на общую концентрацию марганца. |
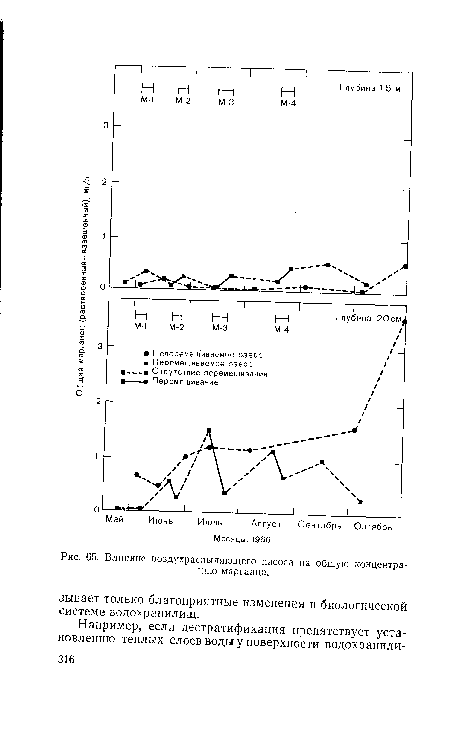 |
Далее
| Количество планктона на глубине 1,5 м в трех исследуемых |
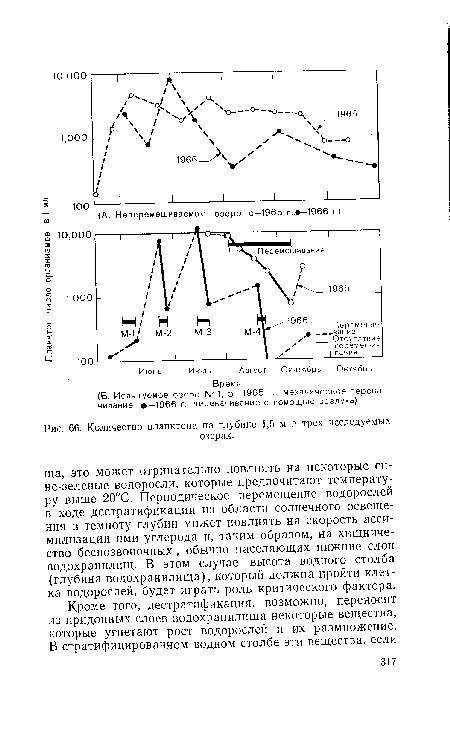 |
Далее
| Состав урожая планктона в перемешиваемом озере в 1965 г. |
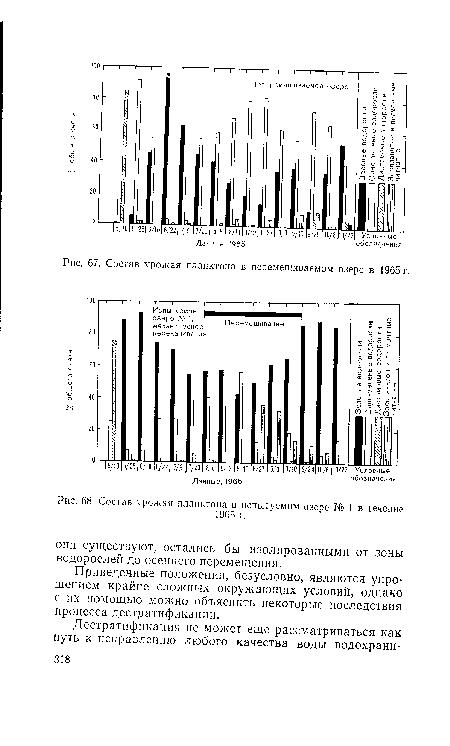 |
Далее
| Состав урожая планктона в испытуемом озере № 1 в течение |
|
Далее
| Состав урожая планктона в перемешиваемом озере в 1966 г. |
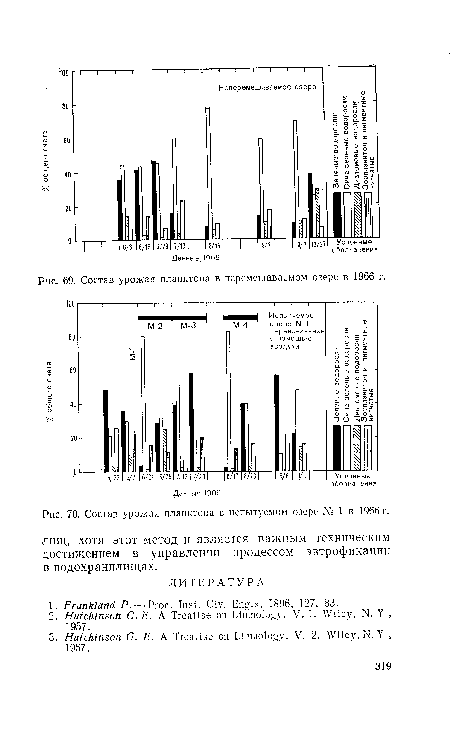 |
Далее
| Состав урожая планктона в испытуемом озере № 1 в 1966 г. |
|
Далее