| И 1.1. Схематическое изображение микрофибриллы природной целлюлозы. |
 |
Далее
| Строение микрофибриллы природной целлюлозы по Мэнли. |
 |
Далее
| Пучок микрофибрилл бактериальной целлюлозы, ассоциированных с более длинными нитями. Отчетливо видно, что отдельные микрофибриллы имеют одинаковую длину, а также разрыхление в середине пучка. |
 |
Далее
| Электронно-микроскопические снимки внутренней ламелли боковой стенки [43]. Ось клетки вертикальна. Следует обратить внимание на две изогнутые микрофибриллы, расположенные под прямыми микрофибриллами. Они не могли быть изогнуты вследствие механического воздействия при получении среза, поскольку они лежат под слоем прямых микрофибрилл. Причиной искривления этих микрофибрилл являются, вероятно, специфические условия их образования. |
![Электронно-микроскопические снимки внутренней ламелли боковой стенки [43]. Ось клетки вертикальна. Следует обратить внимание на две изогнутые микрофибриллы, расположенные под прямыми микрофибриллами. Они не могли быть изогнуты вследствие механического воздействия при получении среза, поскольку они лежат под слоем прямых микрофибрилл. Причиной искривления этих микрофибрилл являются, вероятно, специфические условия их образования.](/static/pngsmall/254604968.png) |
Далее
| Двойное лучепреломление клеточной мембраны культуры A. xylinum, вызванное добавлением холестерина [54]. |
![Двойное лучепреломление клеточной мембраны культуры A. xylinum, вызванное добавлением холестерина [54].](/static/pngsmall/254604976.png) |
Далее
| Концы коротких невыделенных микрофкбрилл бактериальной целлюлозы, оттененные платиноуглеродным порошком. |
 |
Далее
| Понижение содержания сульфатных групп в ацетилцеллюлозе при растворении в уксусной кислоте, содержащей небольшие количества воды [98]. |
![Понижение содержания сульфатных групп в ацетилцеллюлозе при растворении в уксусной кислоте, содержащей небольшие количества воды [98].](/static/pngsmall/254605036.png) |
Далее
| Влияние концентрации едкого натра на содержание азота в цианэтило-вом эфире целлюлозы [38]. Условия реакции |
![Влияние концентрации едкого натра на содержание азота в цианэтило-вом эфире целлюлозы [38]. Условия реакции](/static/pngsmall/254605172.png) |
Далее
| Зависимость содержания азота в цианэтиловом эфире целлюлозы от количества введенного в реакцию акрилонитрила [38]. Условия реакции |
![Зависимость содержания азота в цианэтиловом эфире целлюлозы от количества введенного в реакцию акрилонитрила [38]. Условия реакции](/static/pngsmall/254605174.png) |
Далее
| Угловое распределение интенсивности дифракции рентгеновских лучей для цианэтилировапной хлопковой пряжи, полученное в условиях, при которых пе происходила усадка пряжи [48]. |
![Угловое распределение интенсивности дифракции рентгеновских лучей для цианэтилировапной хлопковой пряжи, полученное в условиях, при которых пе происходила усадка пряжи [48].](/static/pngsmall/254605184.png) |
Далее
| Диэлектрическая проницаемость циаиэтнлировапноп целлюлозы [66]. |
![Диэлектрическая проницаемость циаиэтнлировапноп целлюлозы [66].](/static/pngsmall/254605190.png) |
Далее
| Фактор диэлектрических потерь цианэтилироваиной целлюлозы при |
|
Далее
| Влияние набухания на выход и молекулярный вес привитых цепей привитого сополимера ацетата целлюлозы с полистиролом, полученного методом совместного облучения [30]. |
![Влияние набухания на выход и молекулярный вес привитых цепей привитого сополимера ацетата целлюлозы с полистиролом, полученного методом совместного облучения [30].](/static/pngsmall/254605374.png) |
Далее
| Выход продуктов привитой сополимеризации при прививке акрило-нптрпла к целлюлозной пленке [36]. |
![Выход продуктов привитой сополимеризации при прививке акрило-нптрпла к целлюлозной пленке [36].](/static/pngsmall/254605388.png) |
Далее
| Зависимость разрывной прочности бумаги от содержания привитого полимера [75]. |
![Зависимость разрывной прочности бумаги от содержания привитого полимера [75].](/static/pngsmall/254605402.png) |
Далее
| Спектры ЭПР облученной целлюлозы I. |
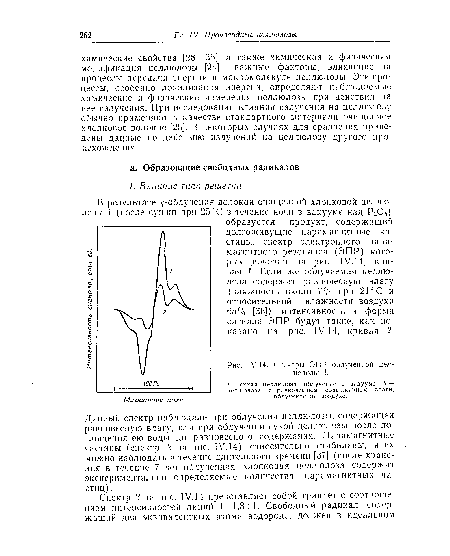 |
Далее
| Спектры ЭПР облученной целлюлозы II. |
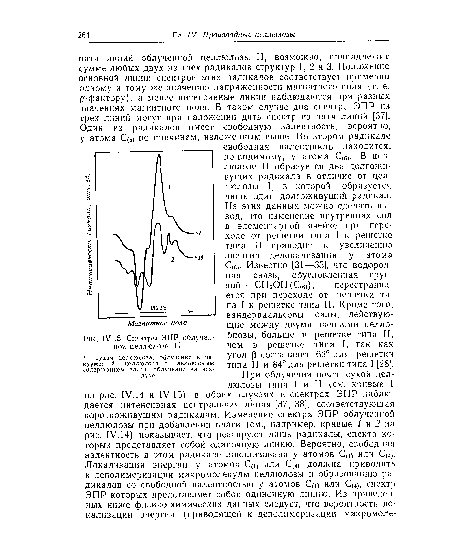 |
Далее
| Л17. Влияние водных растворои хлористого цинка на устойчивость застрявших радикалов в облученной целлюлозе I при 25 °С. |
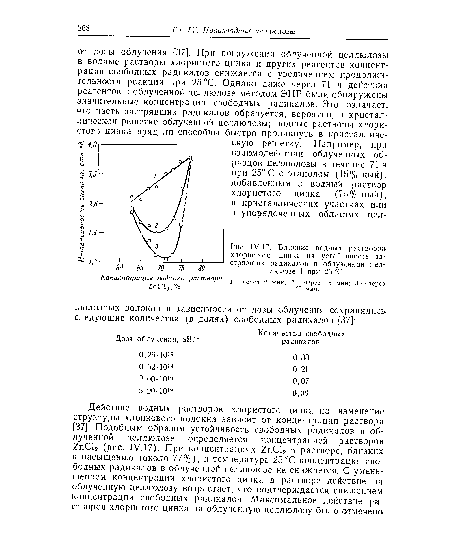 |
Далее
| Изменение спектров ЭПР облученной целлюлозы I в процессе образования привитого сополимера с акрило-нитрилом (15%), растворенным в 75%-ном водном растворе хлористого цинка при 25 °С. Спектры измерены при —100 °С. |
 |
Далее
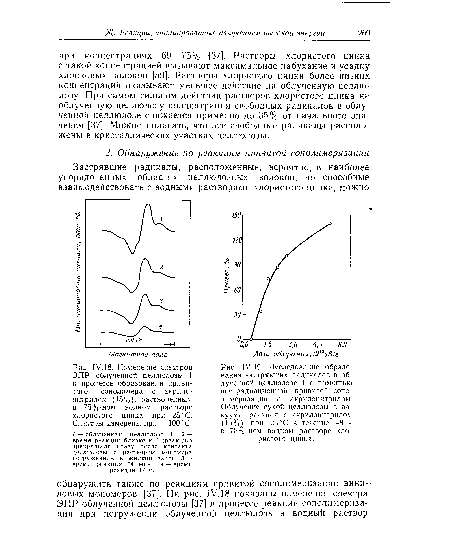
| Исследование образования застрявших радикалов в облученной целлюлозе I с помощью пострадиационной привитой сополимеризации с акрилонитрилом. Облучение сухой целлюлозы в вакууме, реакция с акрилонитрилом (15%) при 25 °С в течение 48 ч в 75%-ном водном растворе хлористого цинка. |
|
Далее
| Поперечный срез прпоитого сополимера хлопкового волокна с акрилопнт-рилом (расстояние между метками 1 мкм). |
 |
Далее
| Поперечный срез облученного хлопка (расстояние между метками 1 мкм). |
 |
Далее
| Влияние процессов передачи и локализации энергии в облученной целлюлозе I на свойства продуктов. |
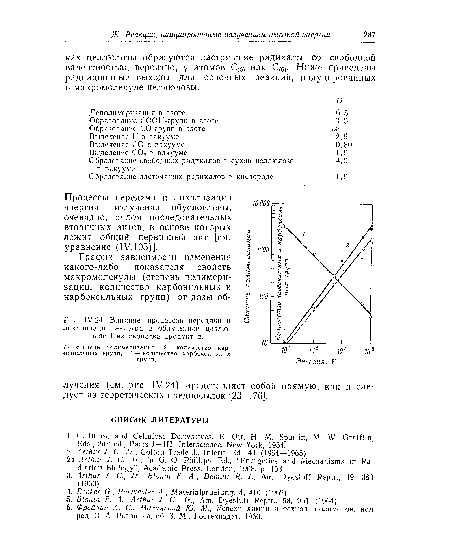 |
Далее
| У.25. Спектры ЭПР радикалов, оознпкающнх при пагревашш целлюлозы и целлобиозы. |
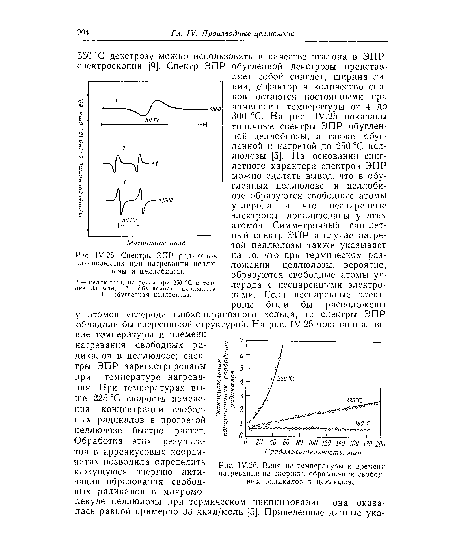 |
Далее
| Влияние температуры и времени нагревания на скорость образования свободных радикалов в целлюлозе. |
|
Далее
| Спектры ЭПР радикалов, генерированных действием света при 25 С. |
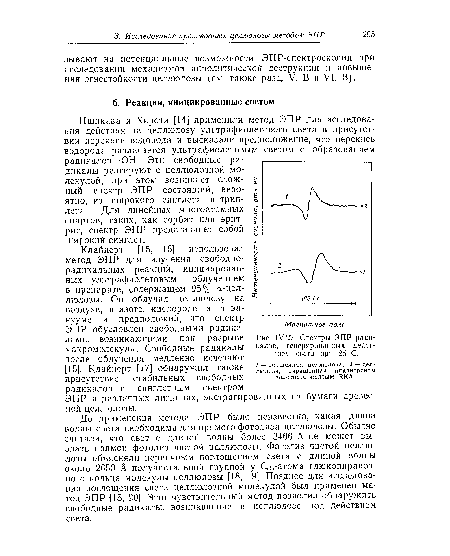 |
Далее
| Спектры ЭПР радикалов, генерированных при реакции иона четырехвалентпого церия с целлюлозой. |
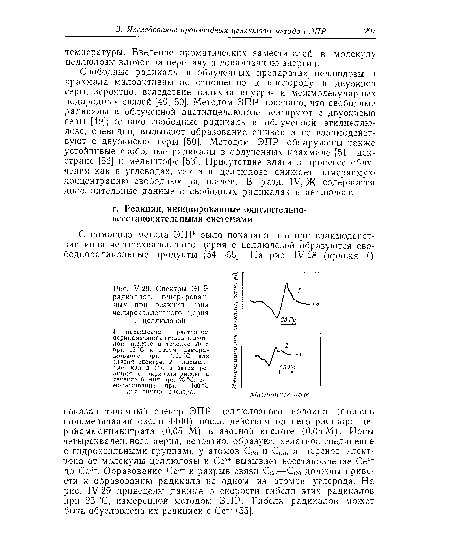 |
Далее
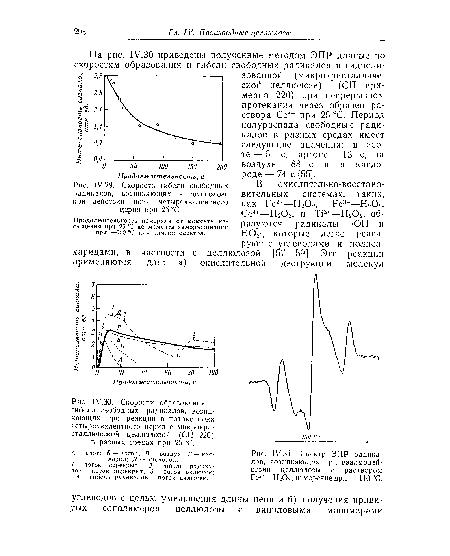
| Спектр ЭПР радикалов, возникающих при взаимодействии целлюлозы с раствором Ре2+—Н202; измерение при —110°С. |
 |
Далее
| Сравнение скорости гидролиза образцов древесной (MS 40 и MS 50) и хлопковой (MS 53 и MS91) целлюлоз в гомогенной среде в концентрированной фосфорной кислоте [6]. |
![Сравнение скорости гидролиза образцов древесной (MS 40 и MS 50) и хлопковой (MS 53 и MS91) целлюлоз в гомогенной среде в концентрированной фосфорной кислоте [6].](/static/pngsmall/254605518.png) |
Далее
| У.З. Соотношение между длиной элементов надмолекулярной структуры, полученных в результате кислотного гидролиза различных образцов целлюлозы в гетерогенной среде, и величиной СП макромолекул [59]. |
![У.З. Соотношение между длиной элементов надмолекулярной структуры, полученных в результате кислотного гидролиза различных образцов целлюлозы в гетерогенной среде, и величиной СП макромолекул [59].](/static/pngsmall/254605528.png) |
Далее
| У.5. Термогравиметрическая кривая а-целлюлозы при пиролизе ее в атмосфере гелия (динамический ТГА) [36]. |
![У.5. Термогравиметрическая кривая а-целлюлозы при пиролизе ее в атмосфере гелия (динамический ТГА) [36].](/static/pngsmall/254605568.png) |
Далее
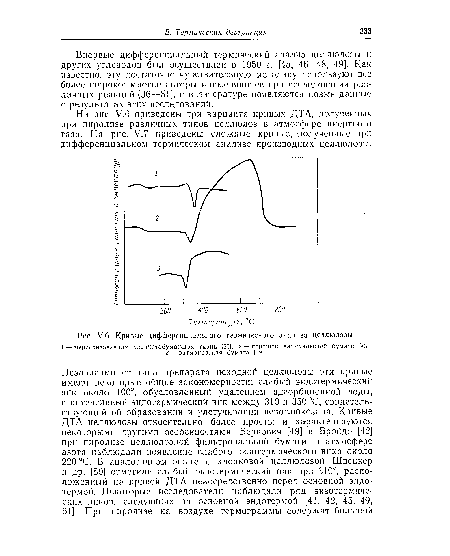
| У.б. Кривые дифференциального термического анализа целлюлозы. |
 |
Далее
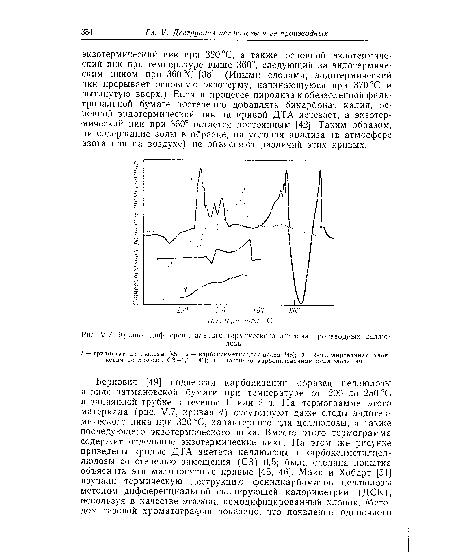
| М.7. Кривые дифференциального термического анализа производных целлюлозы. |
 |
Далее
| Л9. Предполагаемая перегруппировка элементарного звена макромолекулы целлюлозы с образованием 1,2-ангидрида [7]. Ангидрид в открытой форме (А) может перегруппироваться в фурановые производные (см. рис. V. 11). Ангидрид пиранозы в циклической форме (Б) может перегруппироваться в бициклическую |
![Л9. Предполагаемая перегруппировка элементарного звена макромолекулы целлюлозы с образованием 1,2-ангидрида [7]. Ангидрид в открытой форме (А) может перегруппироваться в фурановые производные (см. рис. V. 11). Ангидрид пиранозы в циклической форме (Б) может перегруппироваться в бициклическую](/static/pngsmall/254605604.png) |
Далее
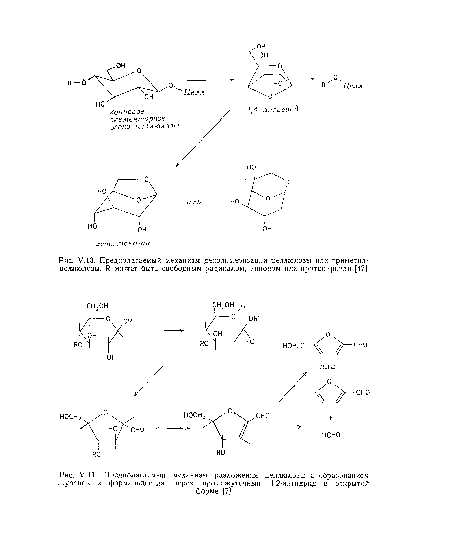
| Предполагаемый механизм разложения целлюлозы с образованием фуранов и формальдегида через промежуточный 1,2-ангидрид в открытой |
 |
Далее
| У.14. Предполагаемый механизм деструкции нитрата целлюлозы [116]. |
![У.14. Предполагаемый механизм деструкции нитрата целлюлозы [116].](/static/pngsmall/254605614.png) |
Далее
| Регулирование образования и активности целлюлазы [35]. |
![Регулирование образования и активности целлюлазы [35].](/static/pngsmall/254605698.png) |
Далее
| Гидролиз хлопковой целлюлозы микроорганизмами и внеклеточными |
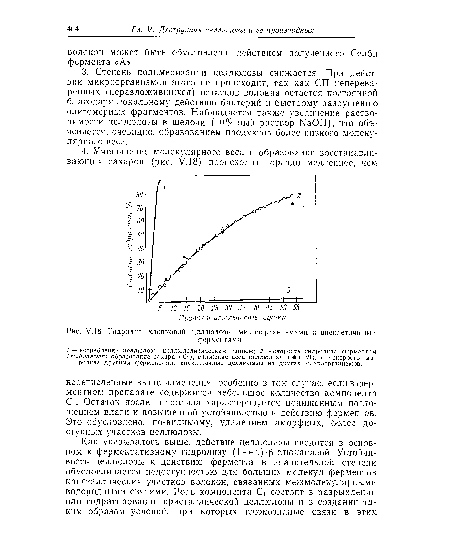 |
Далее
| Микрокристаллы из фибрилл целлюлозы с предельной степенью полимеризации. X 25 ООО. |
 |
Далее
| Микрокристаллы из вискозной кордной нити. Х25 000. |
 |
Далее
| Дополнительное агрегирование микрокристаллов (гель, высушенный |
 |
Далее