| Слоеный пирог» биологии. |
 |
Далее
| Спектр уровней организации. |
 |
Далее
| Схематическое изображение экосистемы пруда. |
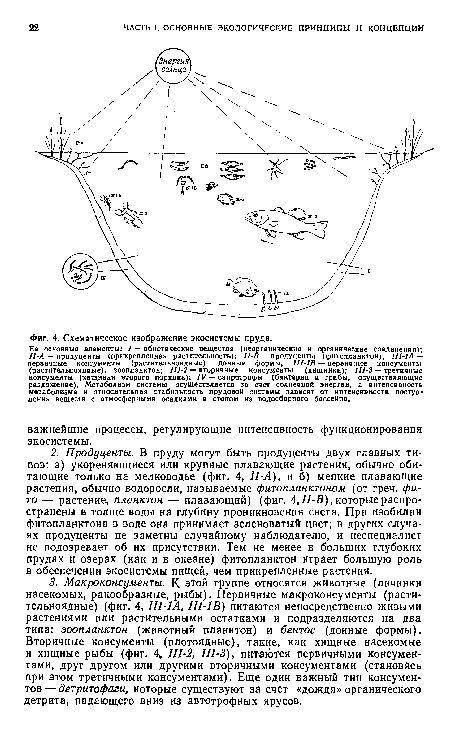 |
Далее
| Измерение метаболизма пруда методом светлых и темных сосудов |
 |
Далее
| Экспериментальный участок водосбора гидрологической лаборатории Ковита в горах на западе Северной Каролины. |
 |
Далее
| Экосистемы луга и залежи. |
 |
Далее
| Примеры упадка экосистем в результате деятельности человека. |
 |
Далее
| Разложение глюкозы бактерией АегоЬаЫег в анаэробных (А) и аэробных (Б) условиях (Пирт, 1957). |
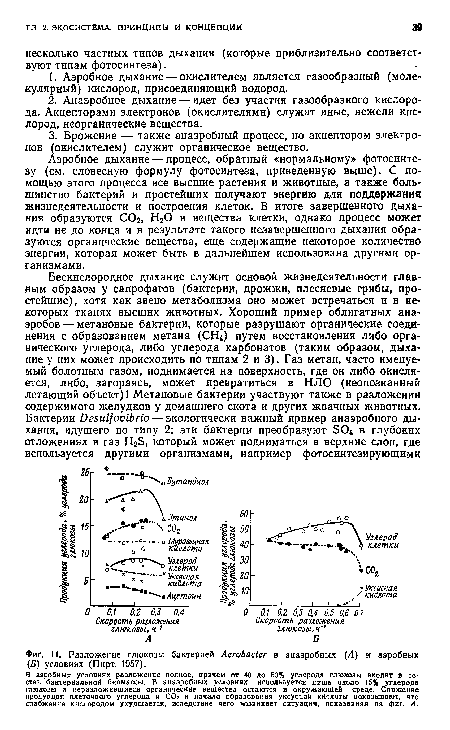 |
Далее
| Три аспекта разложения в экосистеме засоленного марша в Джорджии, где большая часть энергии получается по детритной цепи. |
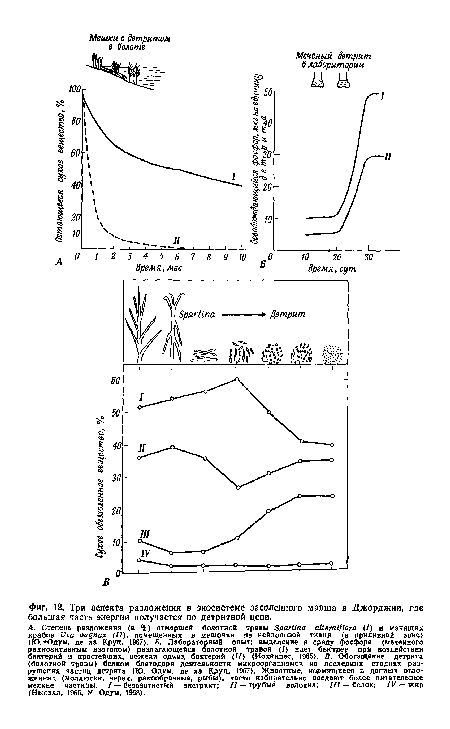 |
Далее
| Элементы кибернетики. |
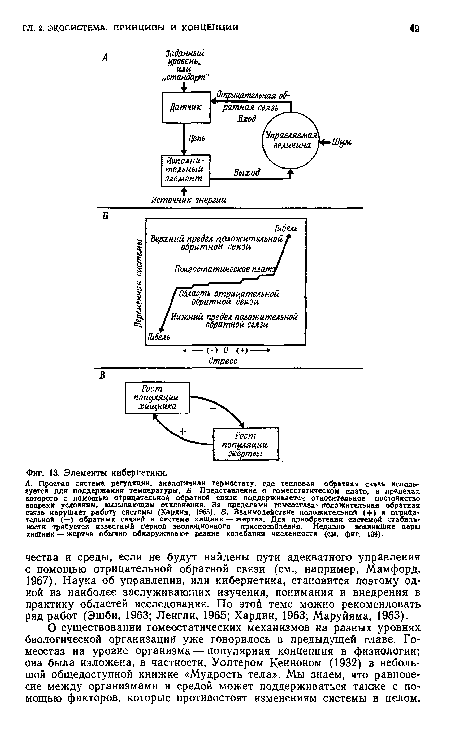 |
Далее
| Диаграмма продуктивности зерновых культур. |
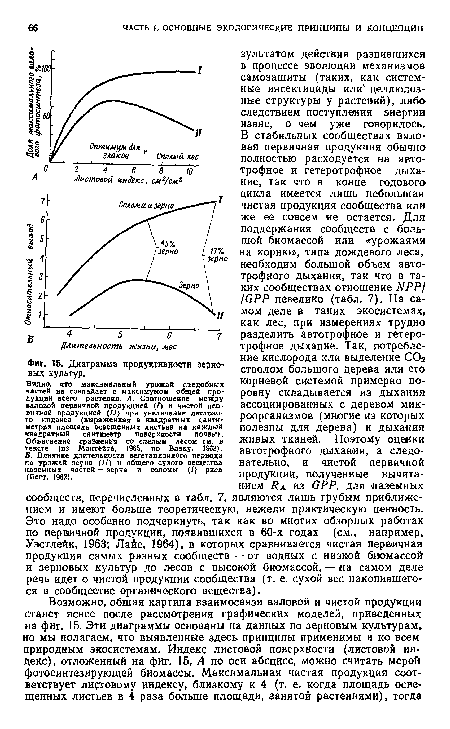 |
Далее
| Мировое распределение первичной продукции экосистем основных типов, выраженное в годовой валовой продукции (в тысячах ккал на 1 м2) (Ю. Одум, 1963). |
 |
Далее
| Упрощенная диаграмма потока энергии, показывающая три tpoфичecкиx уровня (1, 2 н 3) в линейной пищевой цепи (Ю. Одум, 1963). |
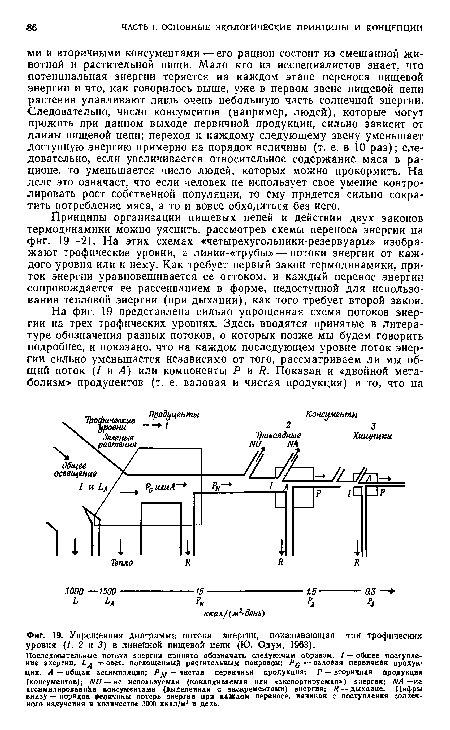 |
Далее
| Диаграмма потока энергии в сообществе с большим «импортом» и меньшим -«экспортом» органического вещества (Г Одум, 1956). |
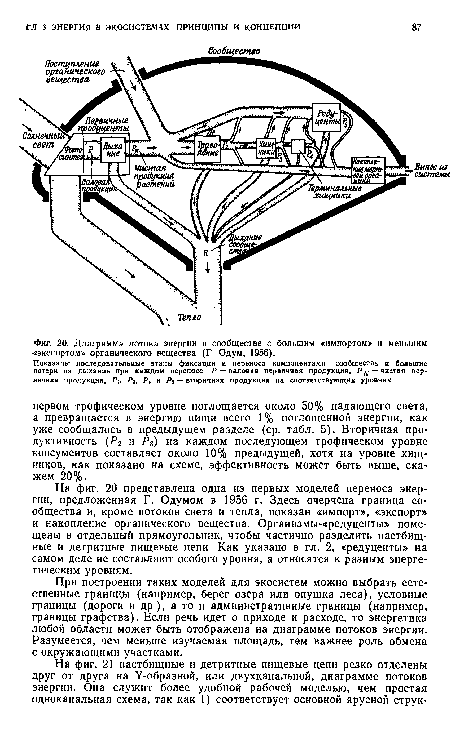 |
Далее
| Компоненты «универсальной» модели экологического потока энергии (Ю. Одум, 1968). |
 |
Далее
| Часть пищевой сети в сообществе небольшого ручья в Южном Уэльса (Джонс, 1949). |
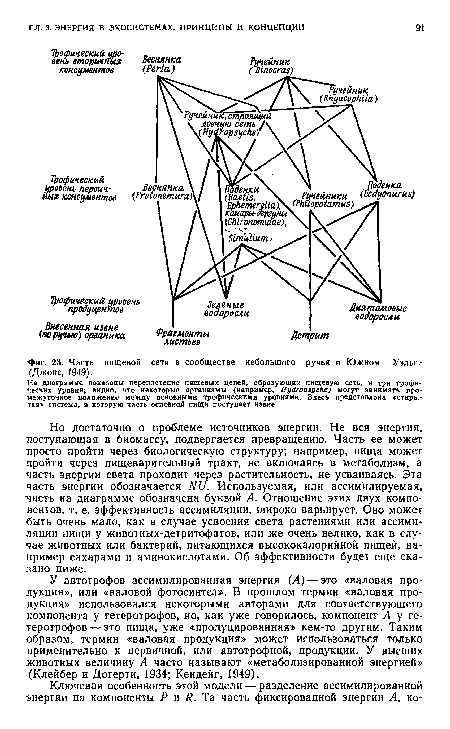 |
Далее
| Схема основных пищевых цепей в пруду (штат Джорджия) для спортивной рыбиой ловли. |
 |
Далее
| Детритная пищевая цепь, базирующаяся на листьях мангровых деревьев на юге Флориды (У. Одум, 1970). |
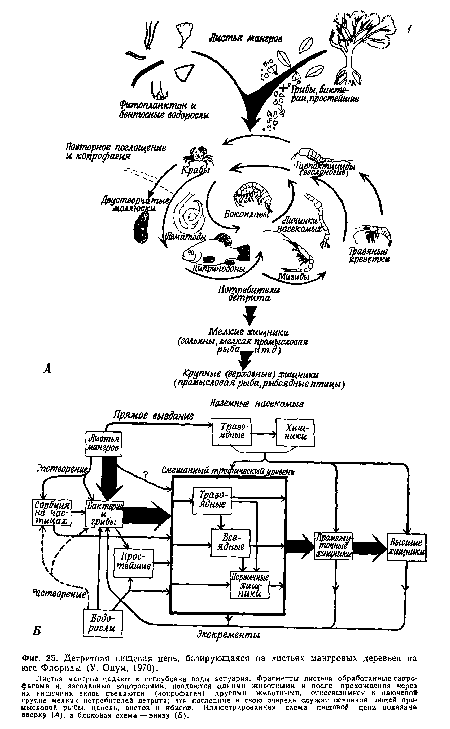 |
Далее
| Связь между эффективностью роста тканей, экологической эффективностью роста и эффективностью ассимиляции в популяциях животных (Уэлч, 1968). |
 |
Далее
| Зависимость между метаболизмом на 1 г биомассы и размерами организма. |
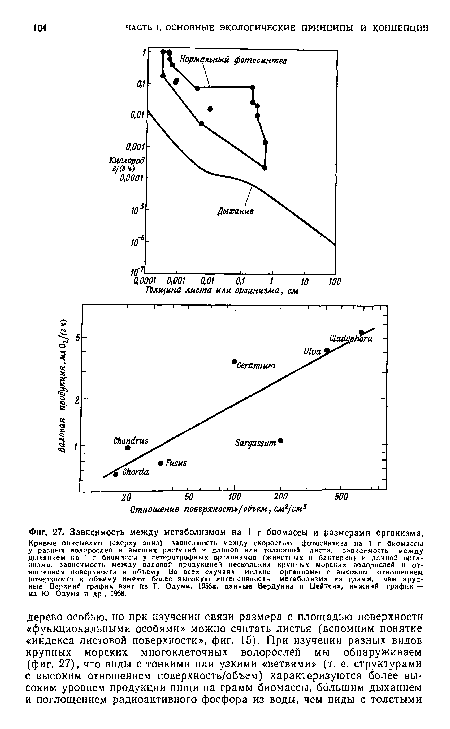 |
Далее
| Экологические пирамиды чисел, биомассы и энергии в разных экосистемах (от открытых вод до больших лесов). |
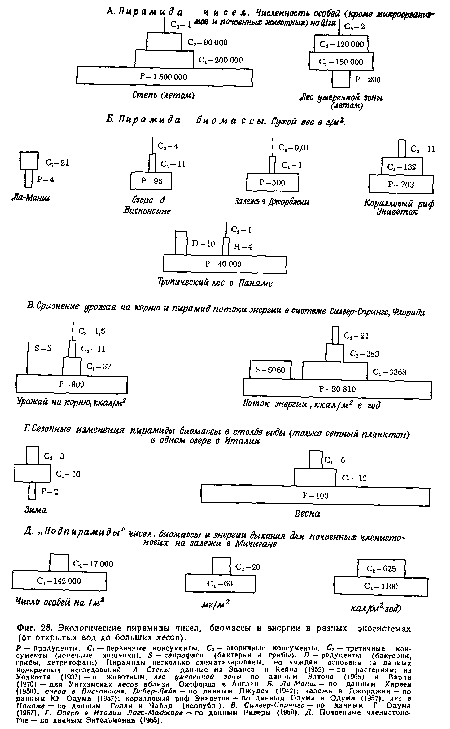 |
Далее
| Три аспекта экосистемы эстуария (Г. Одум. 1969). |
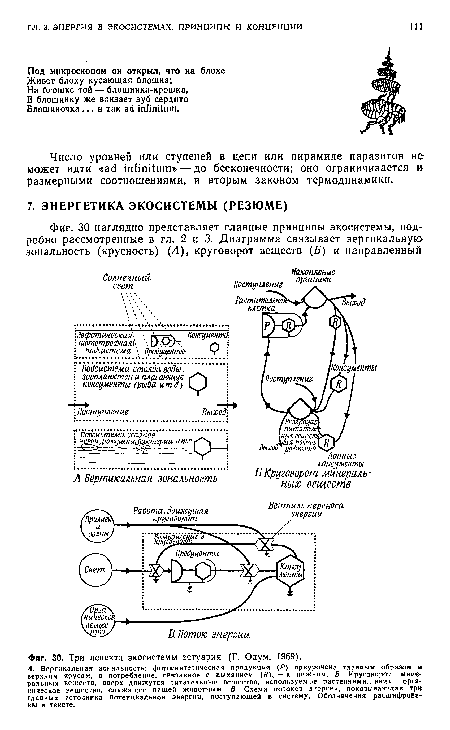 |
Далее
| Распределение дыхания и биомассы (сухой вес) между различными членами бентосного сообщества в маленьком озере на западе Канады (Эффорд, 1969) |
 |
Далее
| Биогеохимический круговорот (заштрихованное кольцо) на фоне упрощен ной схемы потока энергии (Ю. Одум, 1963). |
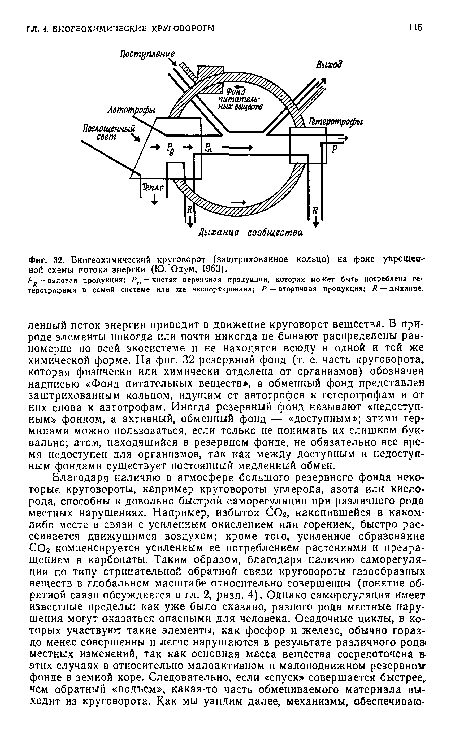 |
Далее
| Два способа изображения биогеохимического круговорота азота. |
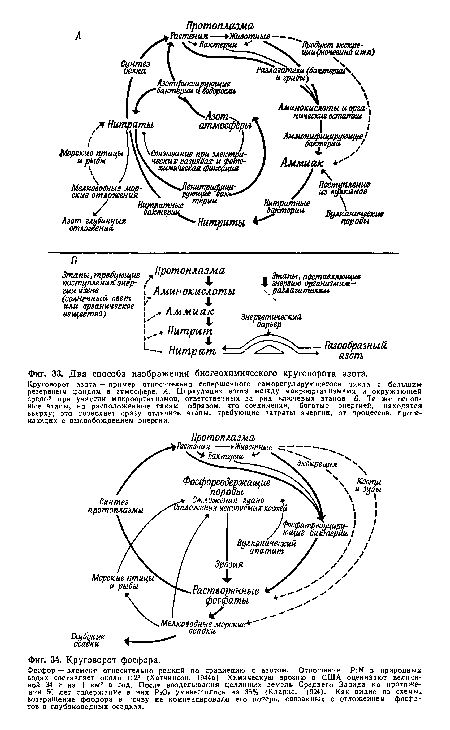 |
Далее
| Круговорот фосфора. |
|
Далее
| Круговорот серы. |
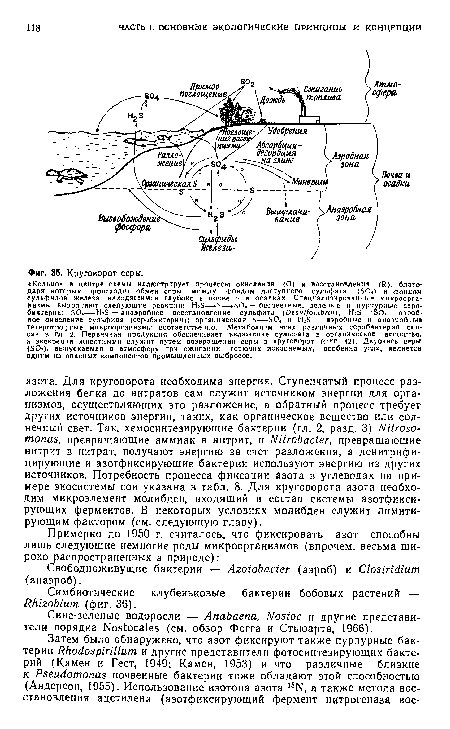 |
Далее
| Корневые клубеньки бобового растения. |
 |
Далее
| Блоковая модель потоков фосфора в экосистеме засоленного марша в штате Джорджия (Помрой и др., 1967). |
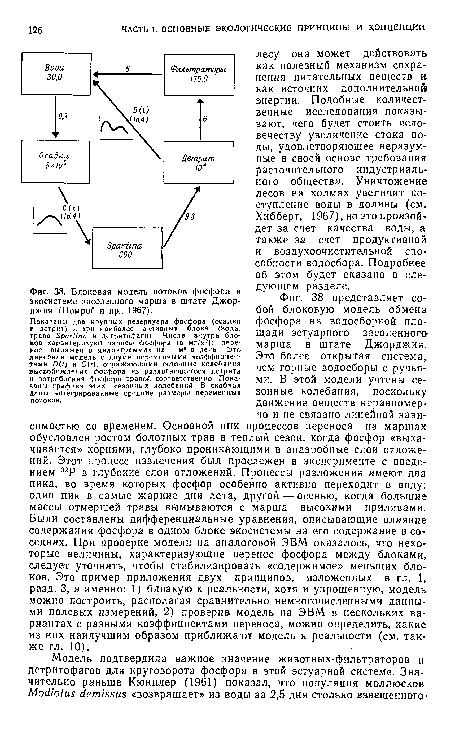 |
Далее
| Схема осадочного цикла. |
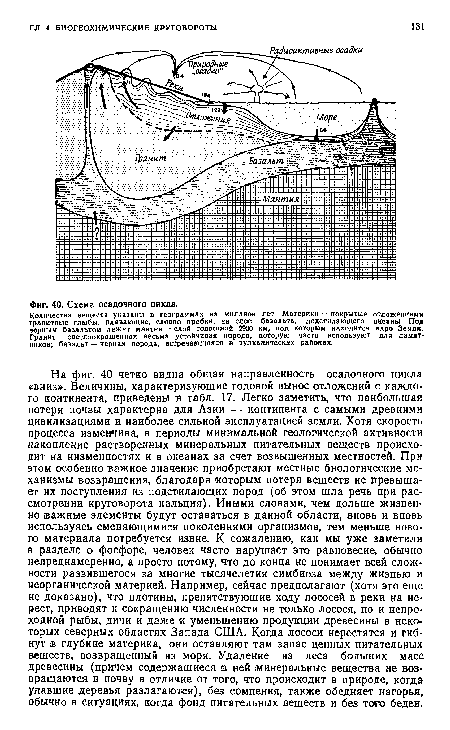 |
Далее
| Распределение органического углерода, накопленного в абиотическом (почва, подстилка) и биотическом (древесина, листья) блоках экосистем северного (/) и тропического (//) леса. |
 |
Далее
| Основные пути возвращения питательных веществ в круговорот. |
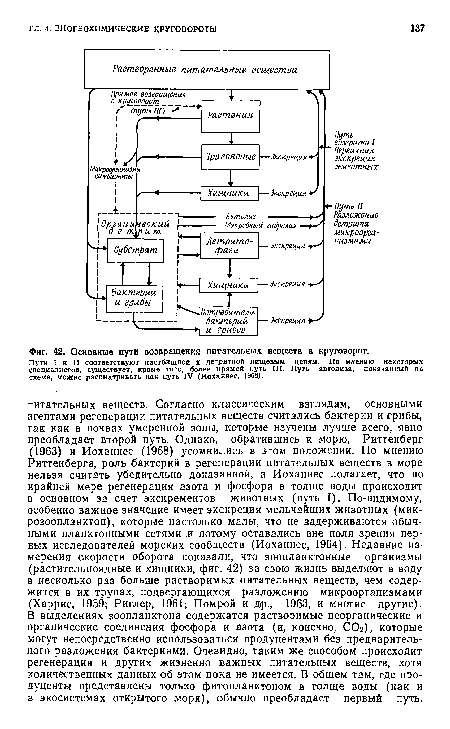 |
Далее
| Сравнение относительных пределов толерантности стенотермных (7 и III) и эвритермных (11) (Рутнер, 1953) организмов. |
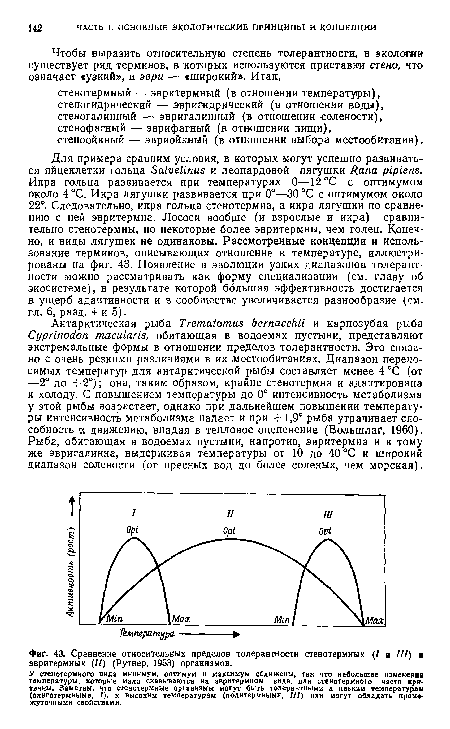 |
Далее
| Температурная компенсация на уровне вида и сообщества. |
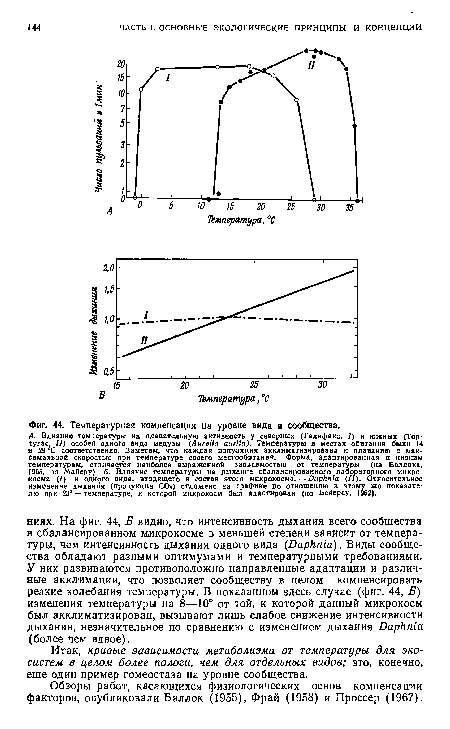 |
Далее
| Модель, обобщающая принципы лими-тирующих факторов (Фрай, 1947). |
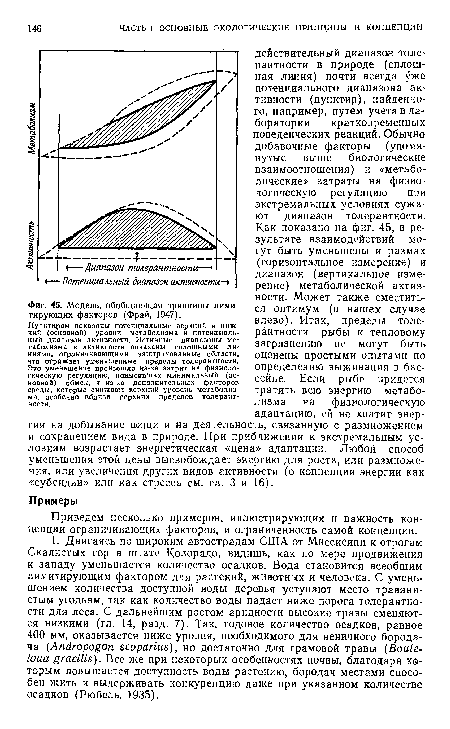 |
Далее
| Метод определения оптимальных районов возделывания сельскохозяйственных культур путем сравнения средних урожаев и изменчивости урожаев из года в год. |
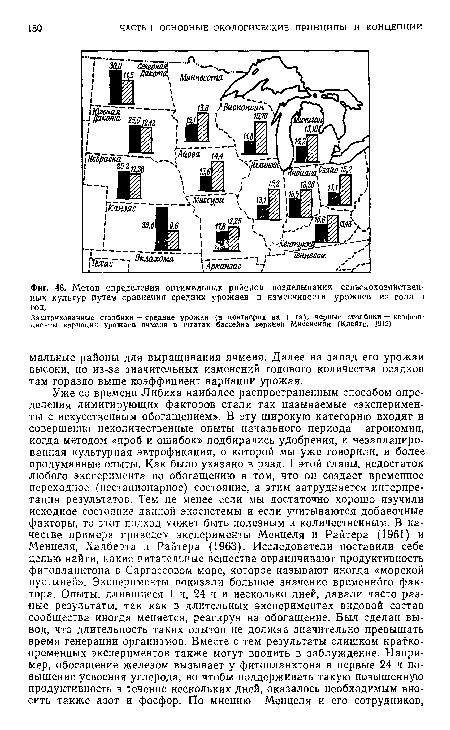 |
Далее
| Управление сроками размножения гольца путем изменении фотопериода (и» Хэзарда и Эддн, 1950). |
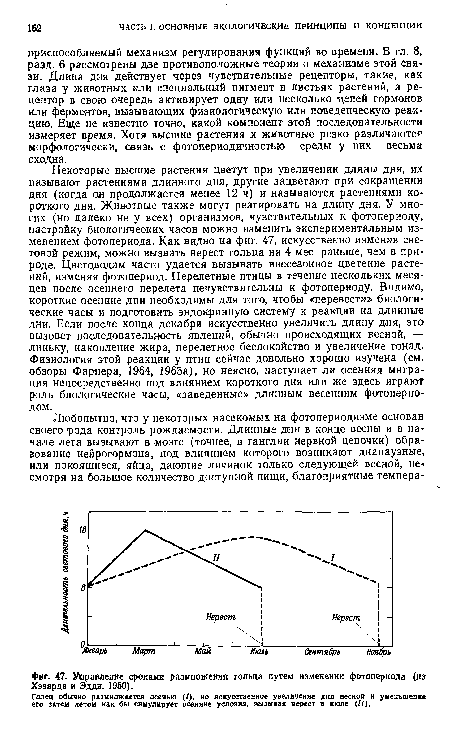 |
Далее
| Электромагнитный спектр и его видимая область (из Колуэлла и др , 1963) |
 |
Далее
| Зависимость фотосинтеза от световой энергии у разных растительных популяций. |
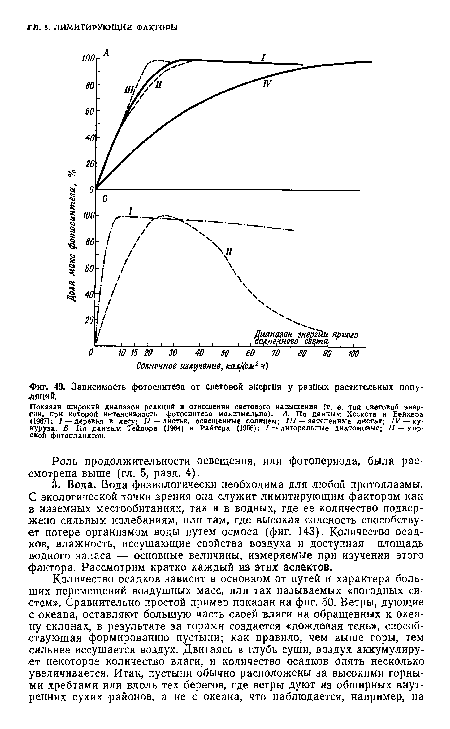 |
Далее
| Годовое количество осадков (вертикальные столбики) в зависимости от высоты местности (бахромчатая линия) по измерениям на ряде станций, тянущихся от Пало-Альто па тихоокеанское побережье к востоку через Береговые хребты н хребет Сьерра-Невада до Оазис-Рэнч в пустыне Невада (по Добенмайру, 1947). |
 |
Далее
| Соотношение между осадками н потенциальной эвапотранспирацией (испарение из почвы + транспирация растений) в трех местообитаниях, соответствующих трем резко различным экологическим областям (Торитуайт, 1955). |
 |
Далее
| Почвенный профиль целины (А) по сравнению с профилем эродированной почвы (Б) в листопадном лесу. |
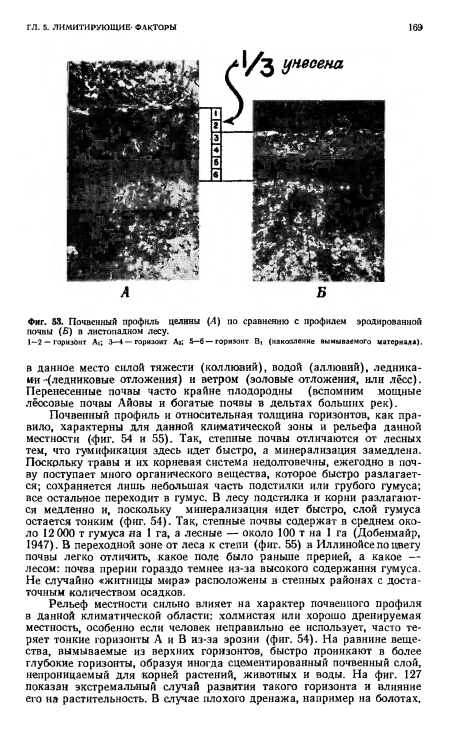 |
Далее
| Упрощенные схемы трех основных типов почвы, характерных для трех биотических зон. |
 |
Далее
| Вересковое болото в Англии, выжженное полосами и пятнами, чтобы увеличить продукцию дичи. |
 |
Далее
| Вертикальный градиент температуры в сильный мороз в центральных районах Аляски |
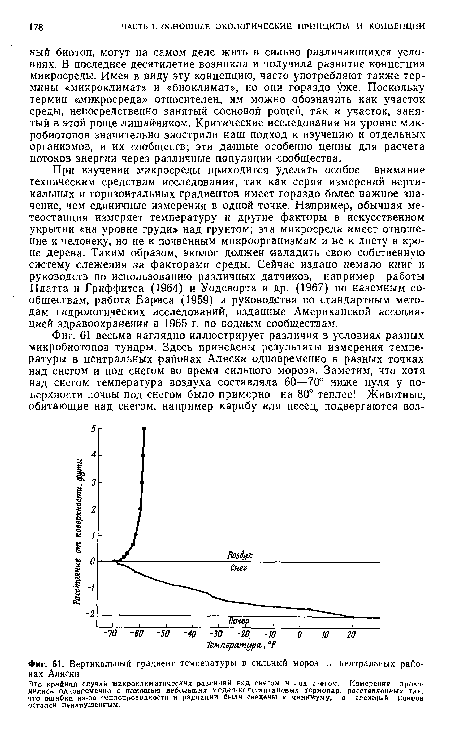 |
Далее
| Солнечное излучение, падающее на равнинную поверхность (//), а также на южный (/) и северный (///) склоны экспериментального водосборного бассейна Ко-внта на западе Северной Каролины. |
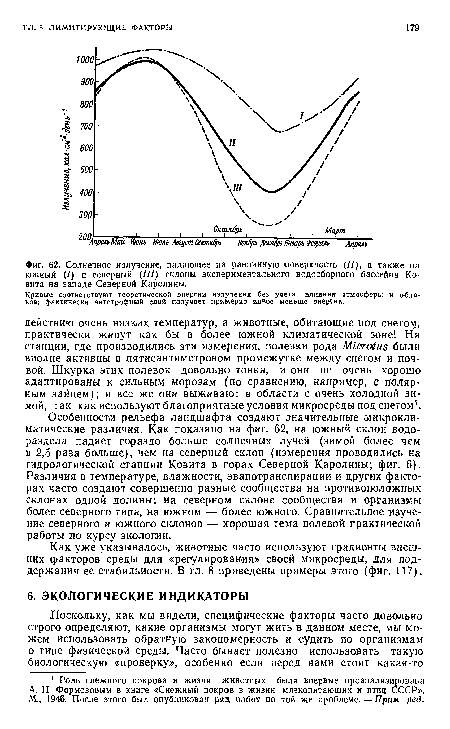 |
Далее
| Распределение популяций доминирующих видов деревьев по гипотетическому градиенту (от 0 до 10), иллюстрирующее распределение популяций в сообществе типа «континуум» (по Уиттэкеру, 1954а) |
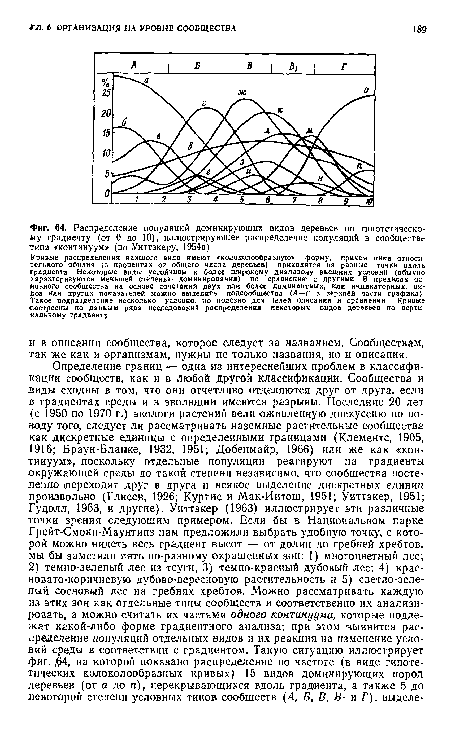 |
Далее
| Градиентный анализ изменения растительности на крутом (/) и пологом (//) склонах (Билз, 1969). |
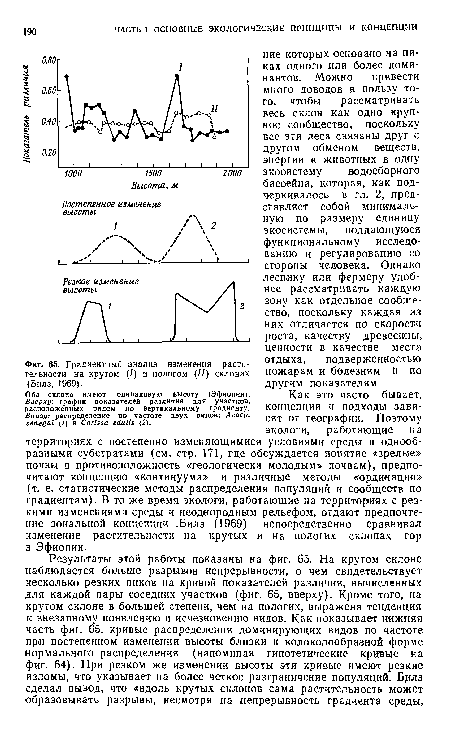 |
Далее
| Использование кривых зависимости между числом видов и их обилием дл$г сравнения видового разнообразия в различных местообитаниях. |
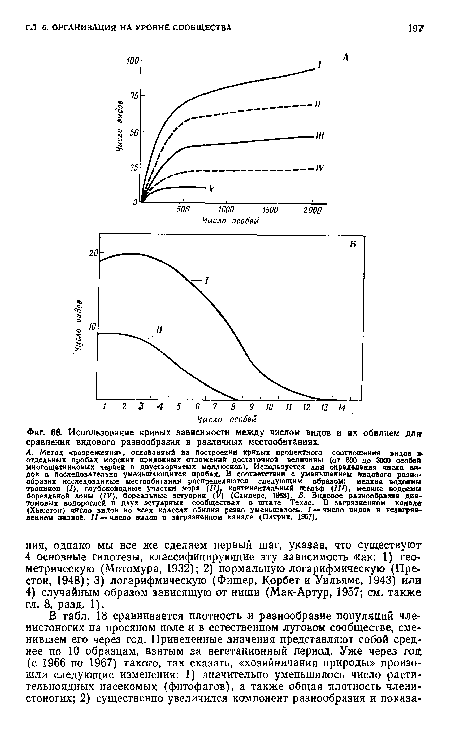 |
Далее
| Вертикальное распределение трех непромысловых видов рыб в середине лета в трех искусственных водоемах в долине Теннесси. |
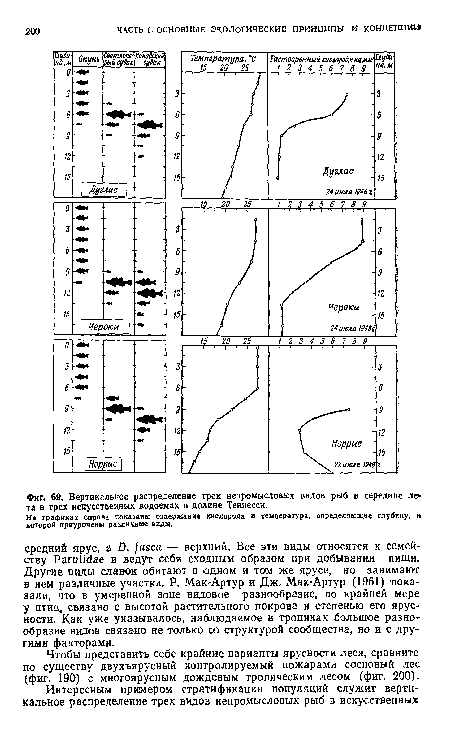 |
Далее
| Реальная пищевая сеть (Б) и две гипотетические сети (А и В). |
 |
Далее
| Распределение пыльцы ископаемых растений в пробах отложений озер на юге Новой Англии (Девис, 1969). |
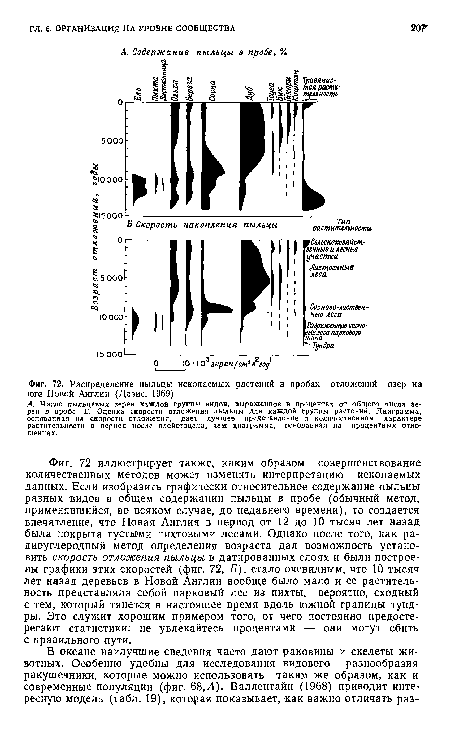 |
Далее
| Диапазон плотности популяций (выраженный в биомассе на 1 га) у раз личных видов млекопитающих в предпочитаемых условиях среды в местностях, где человек не оказывает чрезмерного ограничивающего влияния на численность животных (Мор, 1940). |
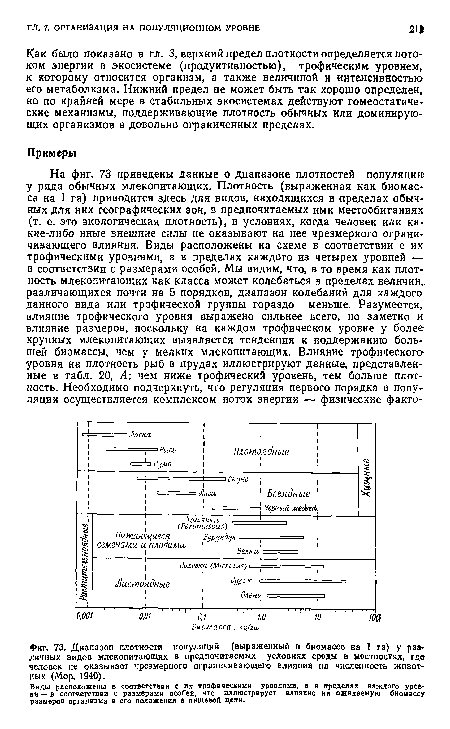 |
Далее
| Кривые роста (вверху) и кривые скорости роста (виизу) популяций двух колоний пчел на одной и той же пасеке (Боденхеймер, 1937). |
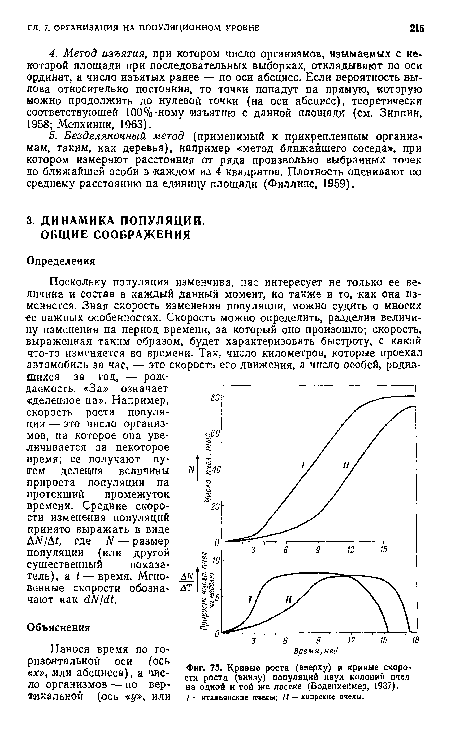 |
Далее
| Возрастные пирамиды. |
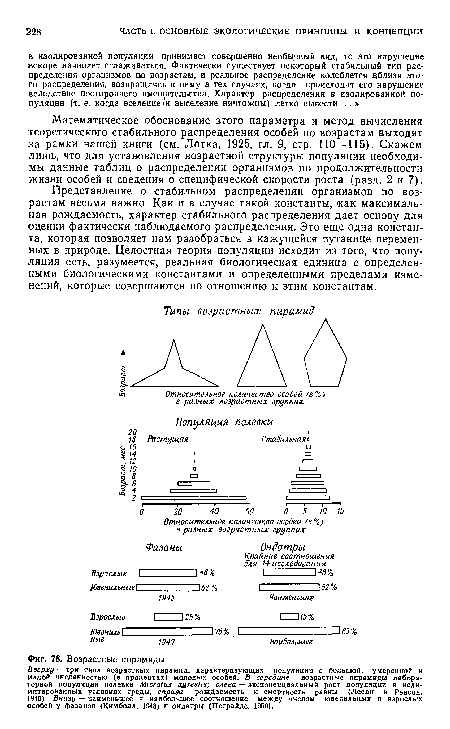 |
Далее
| Доминирование одной возрастной группы у рыб (Хьёрт, 1926). |
 |
Далее
| Возрастные пирамиды человека (для двух местностей в Шотландии) (Дарлинг, 1951). |
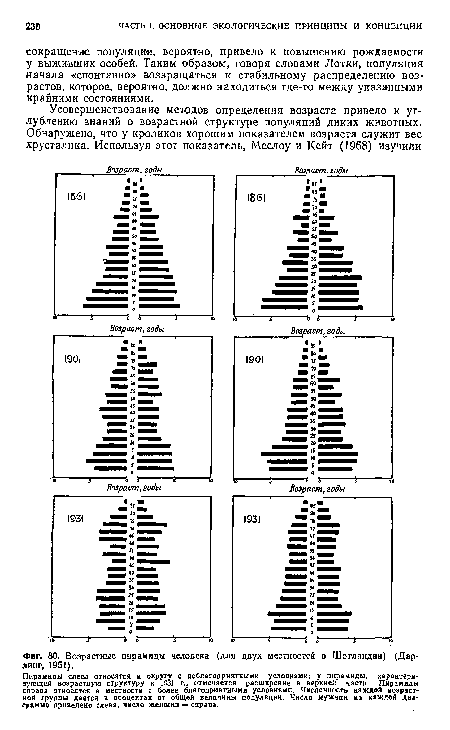 |
Далее
| Кривая роста одной и той же популяции, построенная двумя разными способами. |
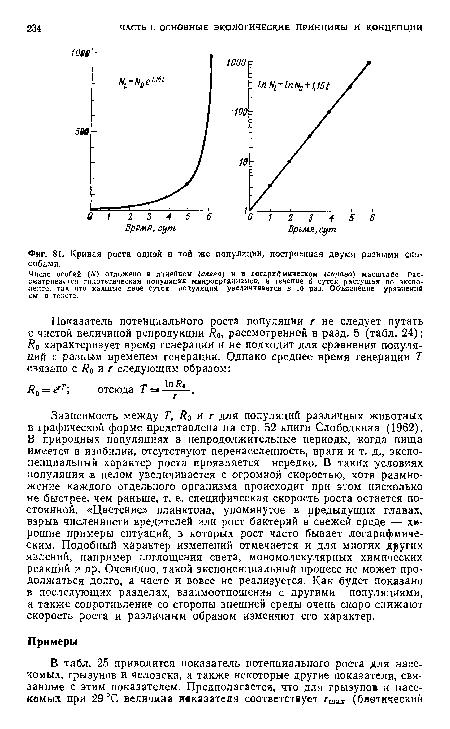 |
Далее
| Некоторые типы кривых роста популяций при изображении в линейном мае- -штабе (Никольсон, 1954). |
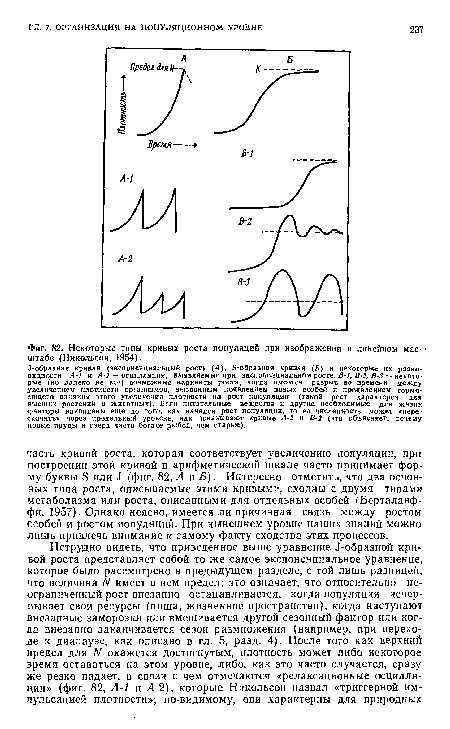 |
Далее
| Рост дрожжей в культуре. |
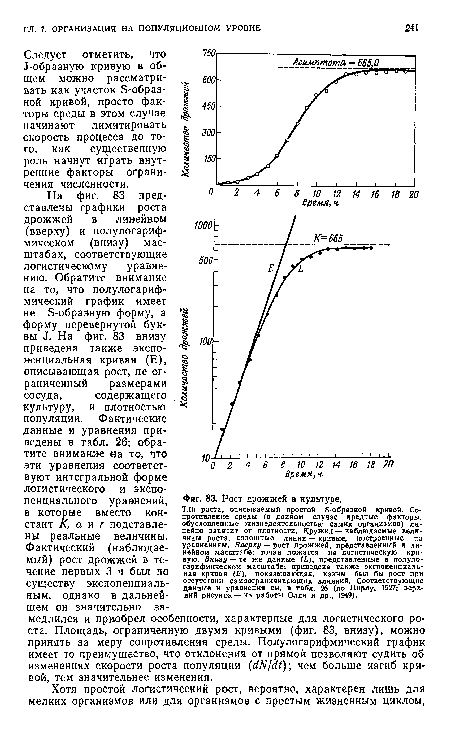 |
Далее
| Сезонные изменения величины популяции взрослых трипсов, живущих на розах (Девидсон и Андреварта, 1948). |
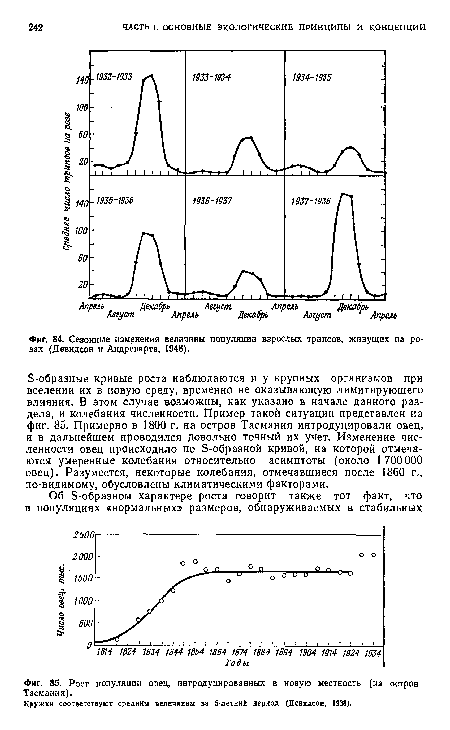 |
Далее
| Рост популяции овец, интродуцированных в новую местность (на остров Тасмания). |
|
Далее
| Сезонные изменения численности семи видов комаров в условиях тропиков в восточной Колумбии (Бейтс, 1945). |
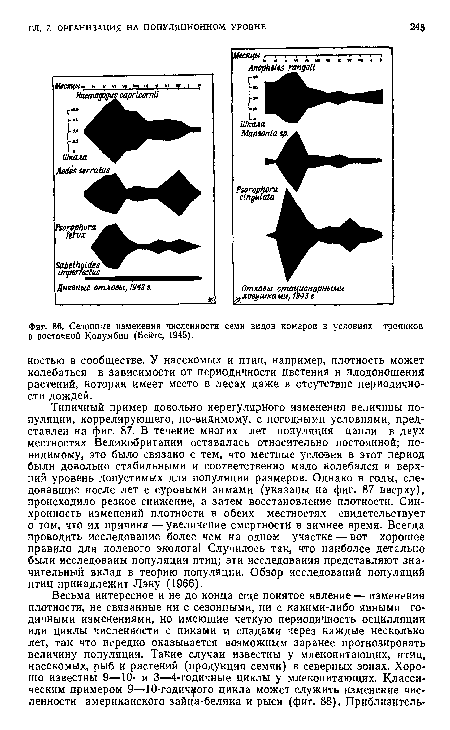 |
Далее
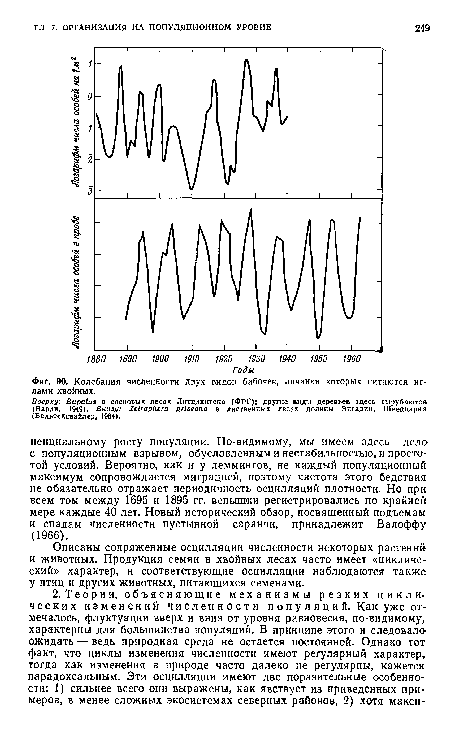
| Американский заяц-беляк, хорошо известный экологам в связи с резко выраженными циклическими изменениями его численности (фиг. 88). |
 |
Далее
| Колебания численности двух видов бабочек, личинки которых питаются иглами хвойных. |
 |
Далее
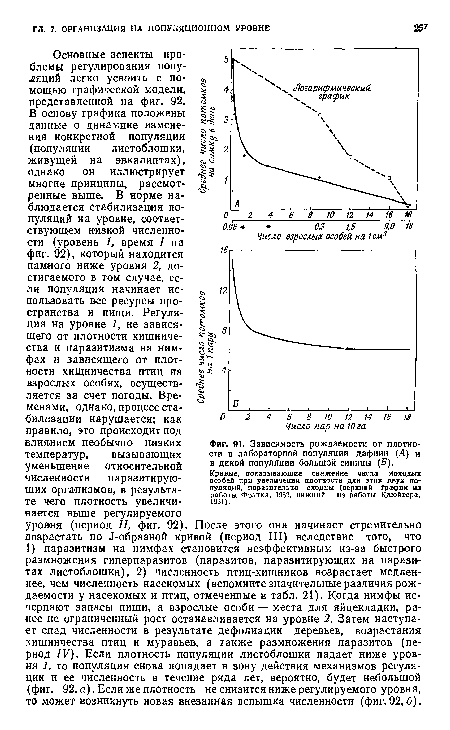
| Зависимость рождаемости от плотности в лабораторной популяции дафнии (Л) и в дикой популяции большой синицы (б). Кривые, показывающие снижение числа молодых особей при увеличении плотности для этих двух популяций, поразительно сходны (верхний график из работы Фрэнка. 1952, нижний — из работы Клюйвера. 1951). |
 |
Далее
| Расселение молодых особей снпухи по отношению к местам гнездований (Стьюарт, 1952). |
 |
Далее
| Годовой бюджет энергии в популяции крапивника, обитающего на засоленном марше, штат Джорджия (Кейл, 1965). |
 |
Далее
| Трн основных типа распределения особей, пар и групп особей в популяции. |
 |
Далее
| Индивидуальные участки и территории (охраняемые индивидуальные участки) у различных животных. |
 |
Далее
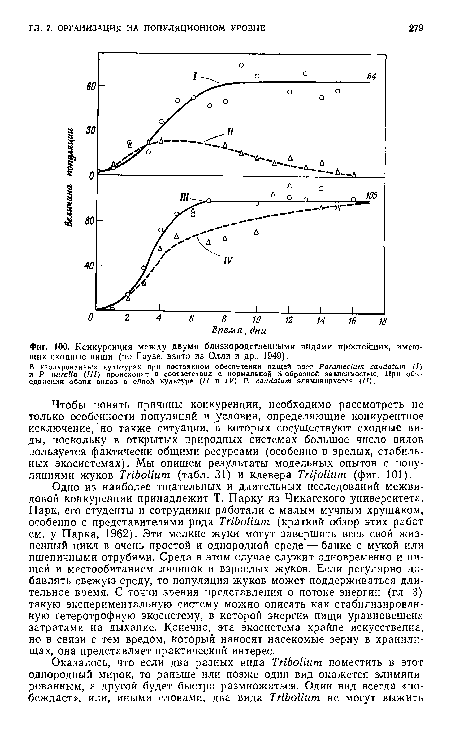
| Конкуренция между двумя близкородственными видами простейших, имеющих сходные ниши (по Гаузе, взято из Олли и др., 1949). |
 |
Далее
| Конкуренция между парами видов дафний в культурах. |
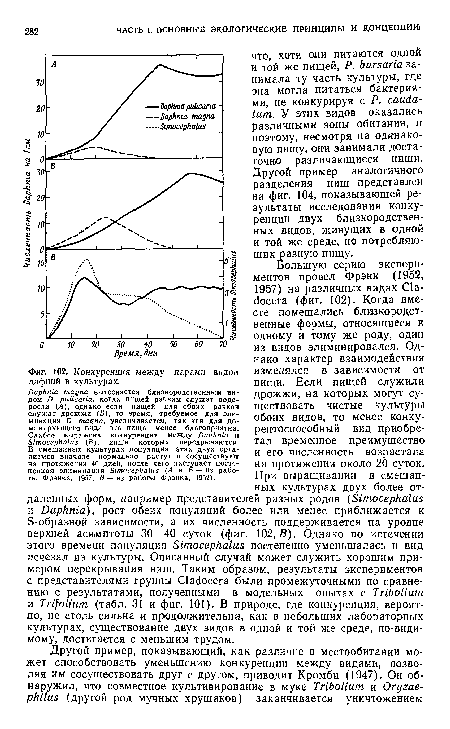 |
Далее
| Влияние конкуренции на распределение местообитаний у птиц (по Свердсо-ну, 1949, с изменениями). |
 |
Далее
| Американские каштаны, пораженные паразитическим грибом (Аппалачские горы, Джорджия) |
 |
Далее
| Биомасса и выход продукции в экспериментальной популяции гуппи (Сил-лимен, 1969). |
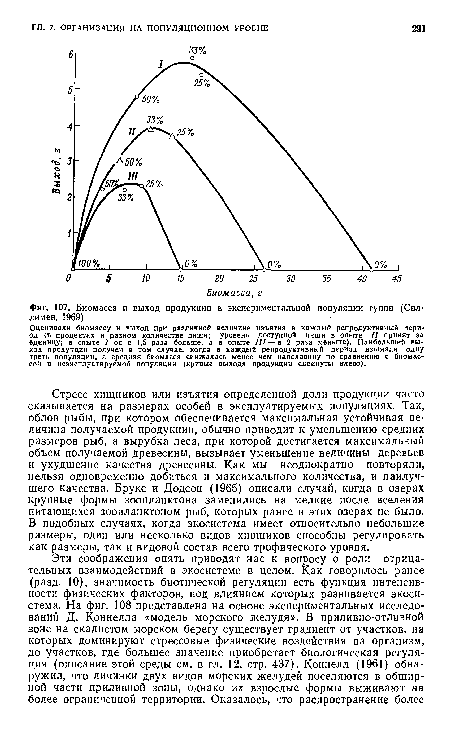 |
Далее
| Факторы, регулирующие распределение двух видов морских желудей в приливной зоне (данные Коннелла, 1961, по Одуму, 1963). |
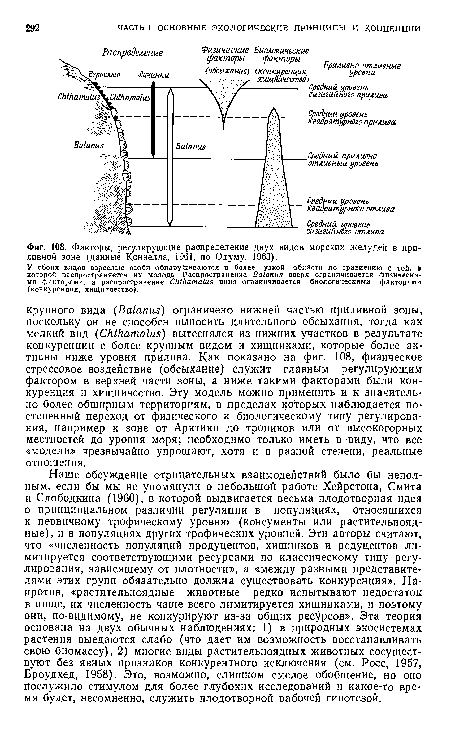 |
Далее
| Уменьшение биомассы тихоокеанской сардины (иваси), подвергавшейся интенсивному облову, и последующее увеличение биомассы ее экологически близкого конкурента — анчоуса, не подвергавшегося эксплуатации (по Силлимену, 1969; данные Мерфи, 1964, и Мерфи и Айзаакса, 1964). |
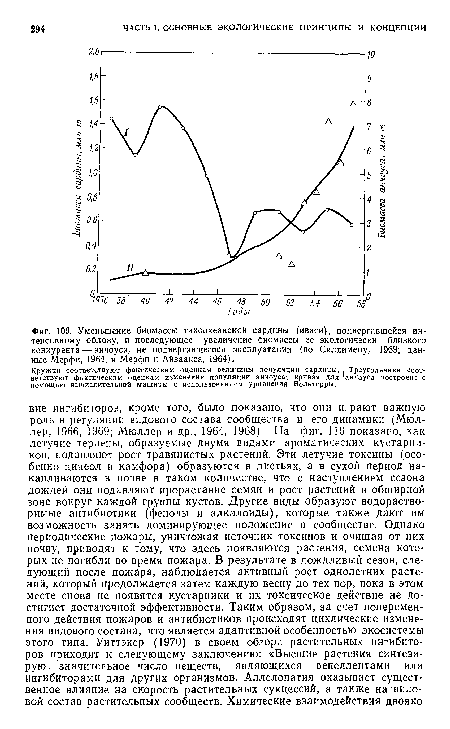 |
Далее
| Типы микоризы. |
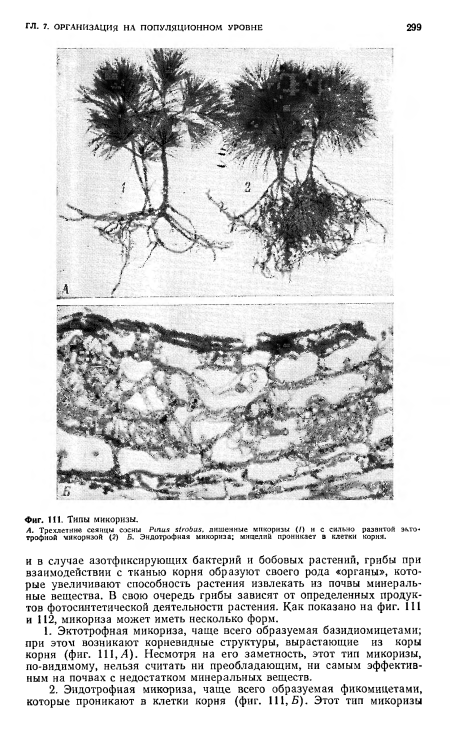 |
Далее
| Направление эволюции от паразитизма к мутуализму у лишайников (Одум, 1963). |
 |
Далее
| Кривые доминирования и разнообразия для гипотетической выборки из сообщества, содержащей 1000 особей 20 видов (из Уиттэкера, 1965). |
 |
Далее
| Два водяных клопа (Hemiptera)—Notonecta (А) и Сопка (Б), которые могут жить в одном и том же местообитании, но занимают разные трофические ниши из-за различий в питании. |
 |
Далее
| Схема климатической ниши кардинала (по Гейтсу, 1969а). |
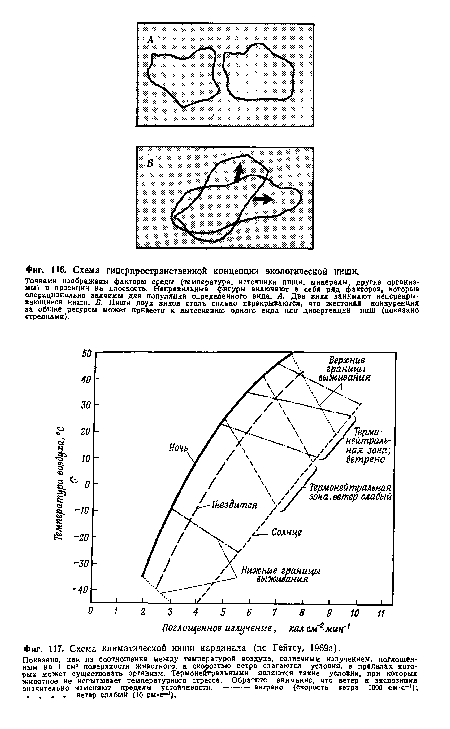 |
Далее
| Пример смещения признаков. |
 |
Далее
| Циркадный ритм летяги (по данным де Курси, 1961). |
 |
Далее
| Относительная важность шести компонентов поведения на разных стадиях филогении (по Детье и Стеллару, 1964). |
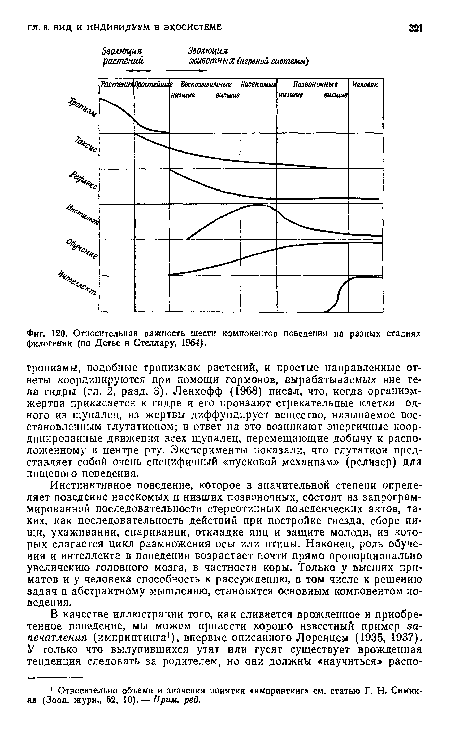 |
Далее
| Положение сообществ различных типов в классификации, основанной на метаболизме (Г. Одум, 1956). |
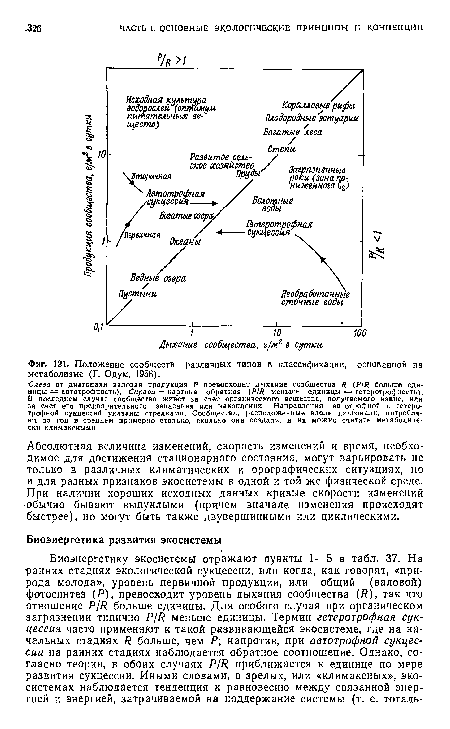 |
Далее
| Сравнение энергетики развития экосистемы леса (Л) и лабораторного микрокосма (£). |
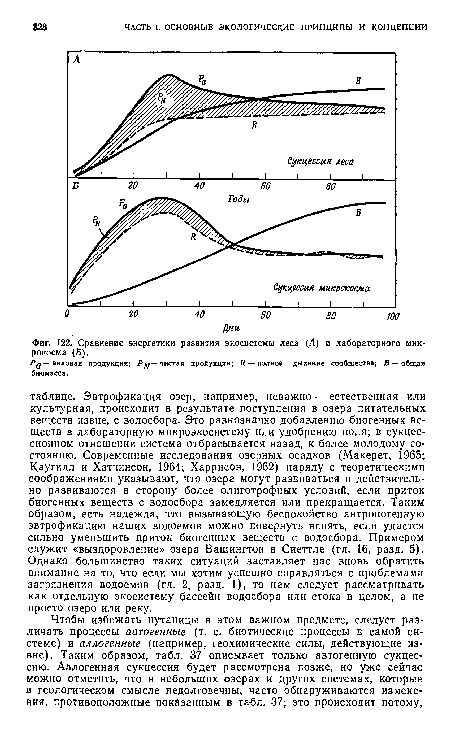 |
Далее
| Разнообразие по обилию видов в четырех ярусах лесной сукцессии в районе Пидмонта (Джорджия). |
 |
Далее
| Вторичная сукцессия в районе Пидмонта, на юго-востоке США. |
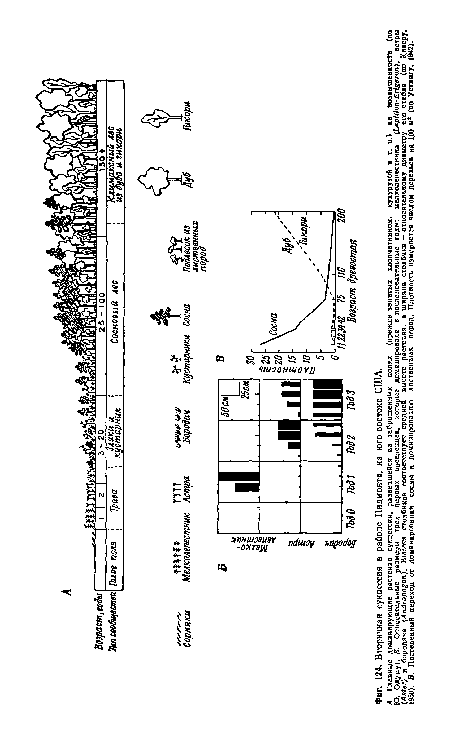 |
Далее
| Сукцессия в культуре сенного отвара со сменой доминирующих видов. Это лабораторный пример гетеротрофной сукцессии. |
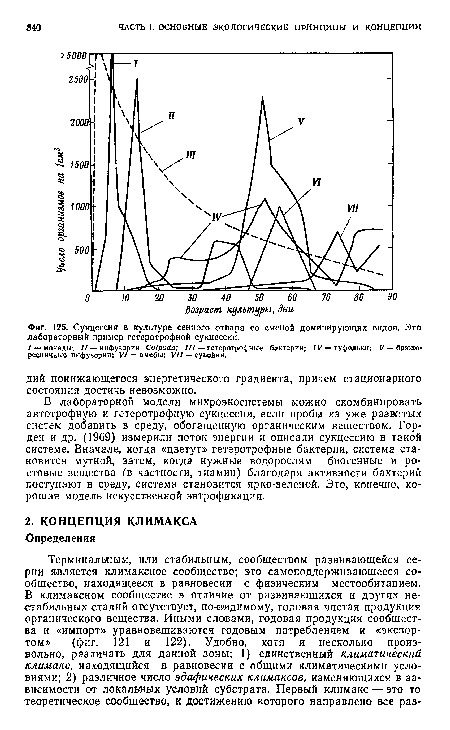 |
Далее
| Блоковая модель основных необходимых человеку типов среды обитания, расчлененных в соответствии с потребностями развития экосистемы и ресурсов жизненного цикла (объяснения в тексте). |
 |
Далее
| Эволюция биосферы и ее кислородной атмосферы (объяснения в тексте). |
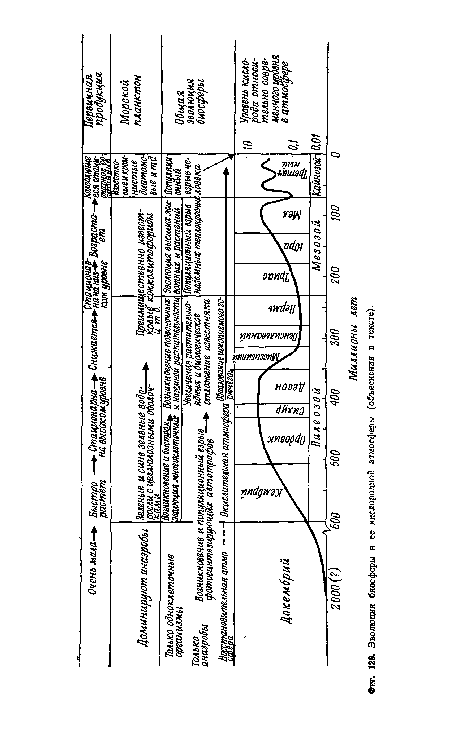 |
Далее
| Различные пути изменения видовых популяций, приводящие к эволюционным изменениям в сообществе |
 |
Далее
| Процессы и структуры в экологических системах можно представить в виде «черных ящиков», состоящих из более простых черных ящиков и образующих иерархию сложности. |
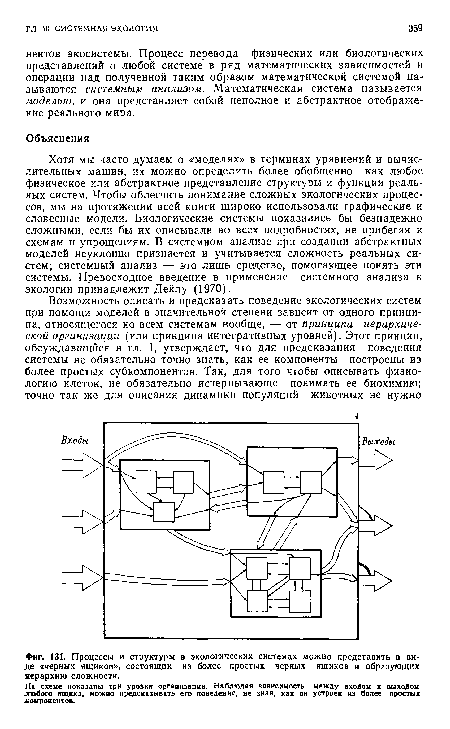 |
Далее
| Основная система обратной связи с точки зрения теории регулирования (по Уотермену, 1968). |
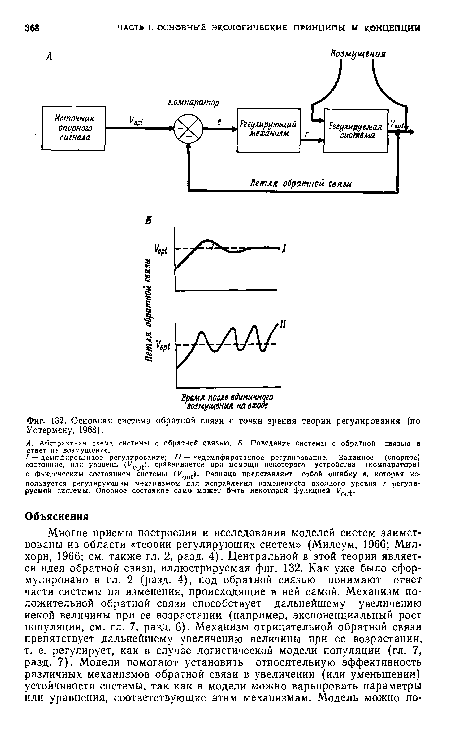 |
Далее
| Экспериментальная блоковая модель взаимодействия паразит—хозяин (Холдинг и Юинг, 1969). |
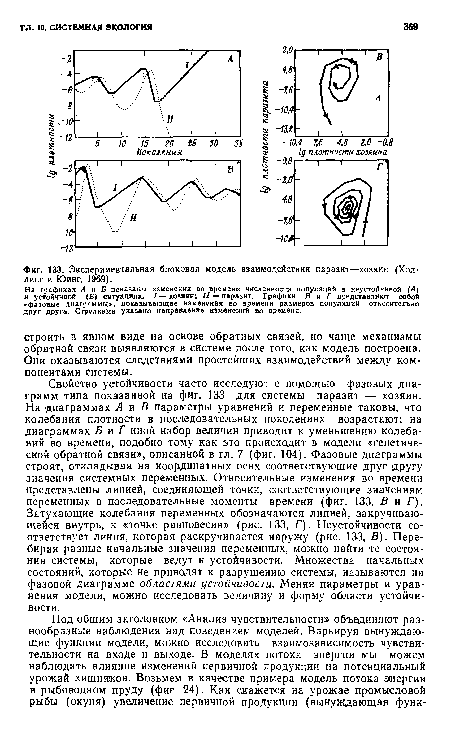 |
Далее
| Блоковая модель изменения биомассы в экосистеме степи (из ван Дайна, 1969). |
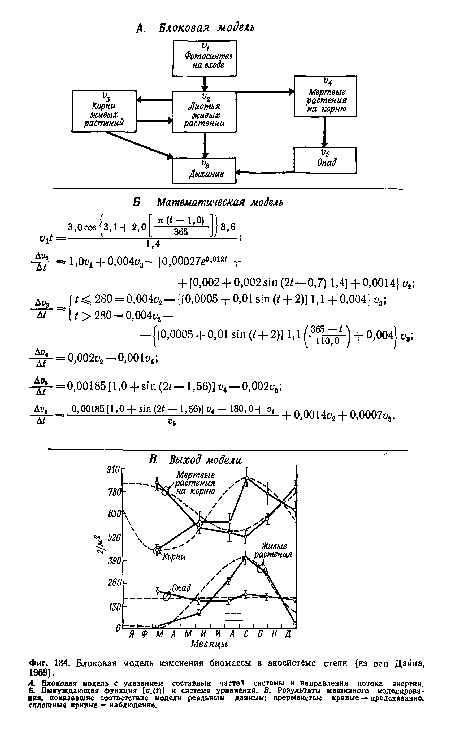 |
Далее
| Крупномасштабная имитационная модель степной экосистемы (нз ван Дайна, 1969). |
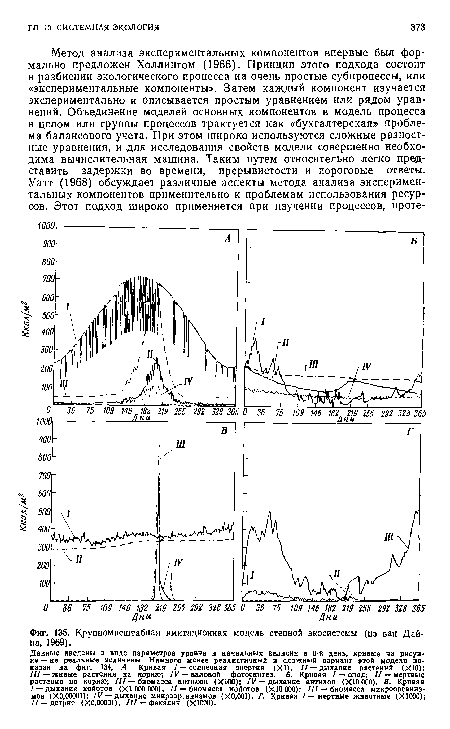 |
Далее
| Имитация растительной сукцессии, последовавшей за жестоким пожаром в окультуренной области, в которой не было крупных травоядных. |
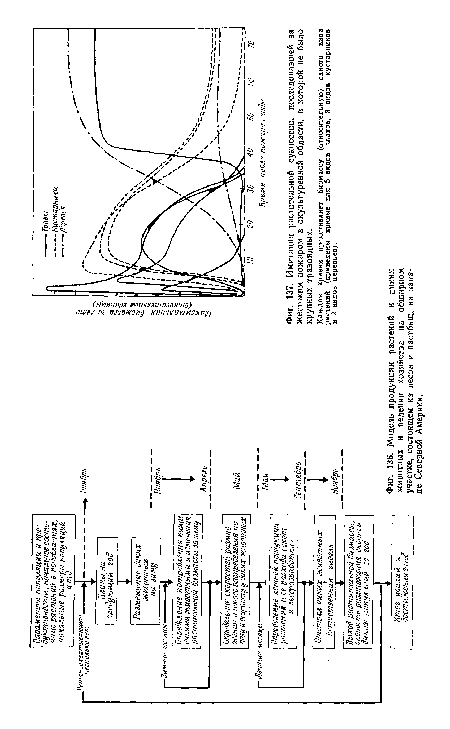 |
Далее
| Модель продукции растений и диких животных и ведения хозяйства на обширном участке, состоящем из лесов и пастбищ, на западе Северной Америки. |
|
Далее
| Пример аналоговой модели в виде электрической цепи (по Одуму, 1967 1967а). |
 |
Далее
| Типы стоячих водоемов. |
 |
Далее
| Две фотографии, иллюстрирующие тесную взаимозависимость проточных водоемов и прилегающих участков суши. |
 |
Далее
| Осморегуляция у пресноводных (Л) и морских (Б) костистых рыб (Флор-кин и Моргулнс, 1949, по Болдуину). |
 |
Далее
| Три главные зоны в озере. |
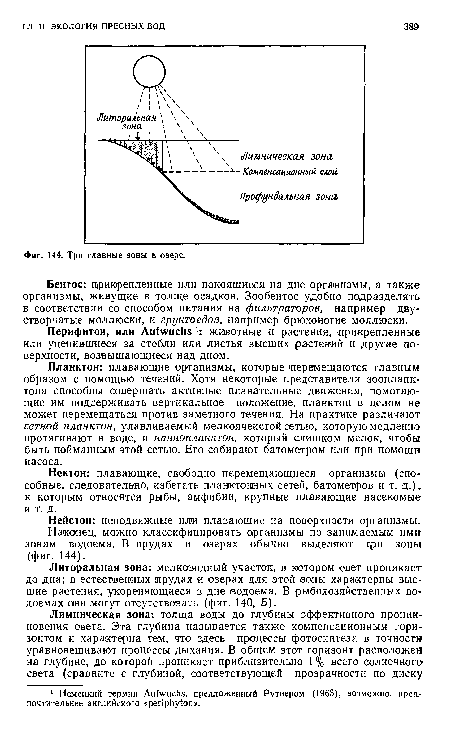 |
Далее
| Некоторые продуценты лентических сообществ; прибрежные растения, укореняющиеся в дне водоема (1—7), нитчатые водоросли (8—9) и фитопланктон (10—20). |
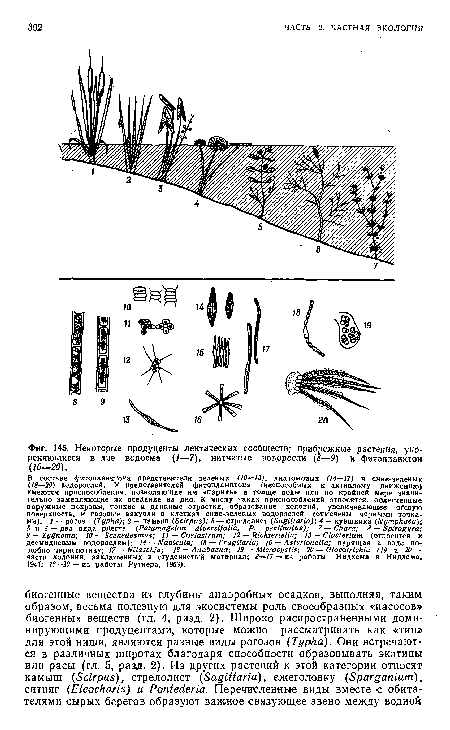 |
Далее
| Типичные представители фауны литоральной зоны прудов и озер (Р. Пен-нак, 1953). |
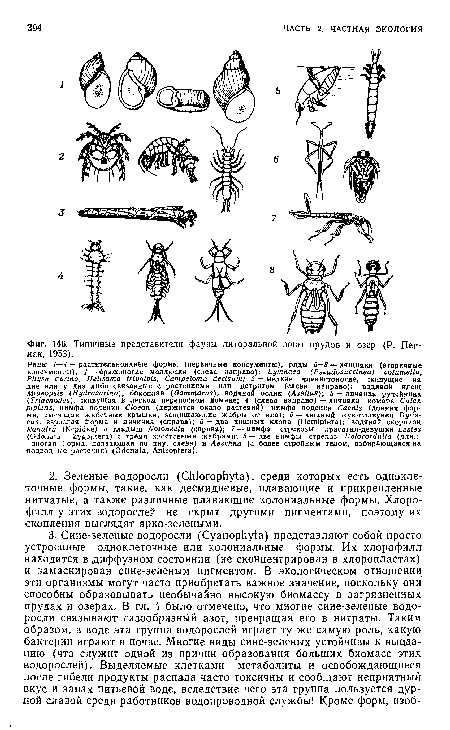 |
Далее
| Различные пресноводные животные (Р. Пеннак, 1953). |
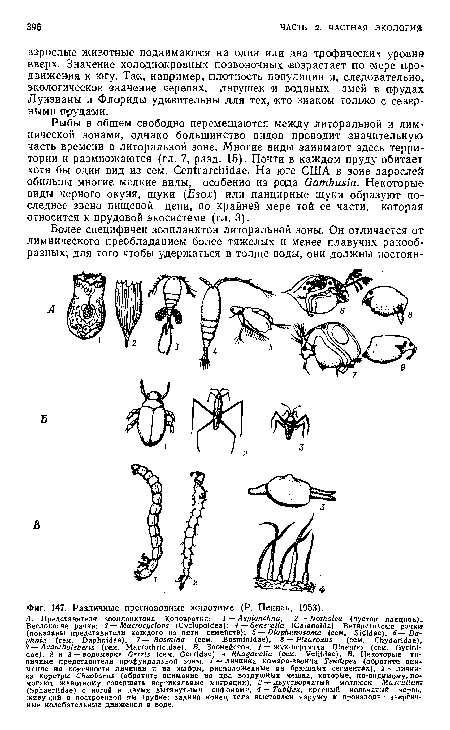 |
Далее
| Возможный механизм вспышки размножения фитопланктона в прудах и озерах умеренной зоны (объяснения в тексте). |
 |
Далее
| Вертикальный ход августовских температур в двух озерах штата Монтана (Райт, 1967). |
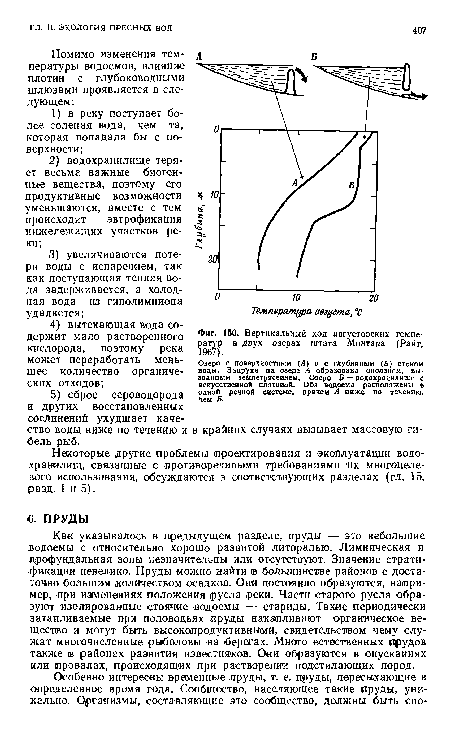 |
Далее
| Сукцессия животных в одном из временных прудов штата Иллинойс (Уэлч, по Шелфорду, 1919). |
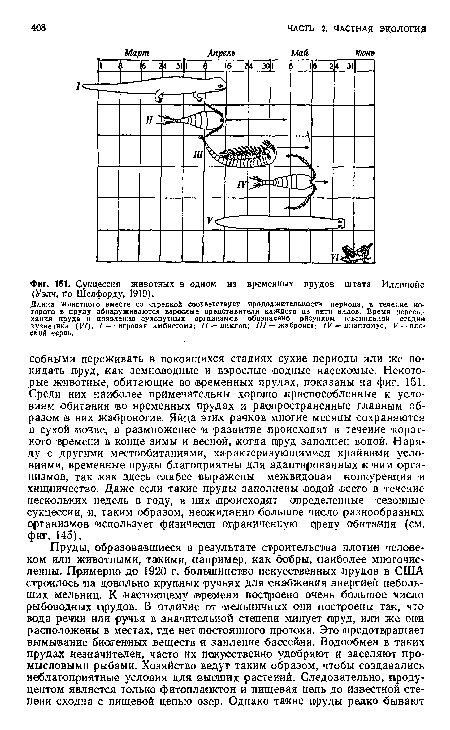 |
Далее
| Гндроклимаграммы температура — соленость для лиманных (солоноватых) и морских районов (Хеджпет, 1951). |
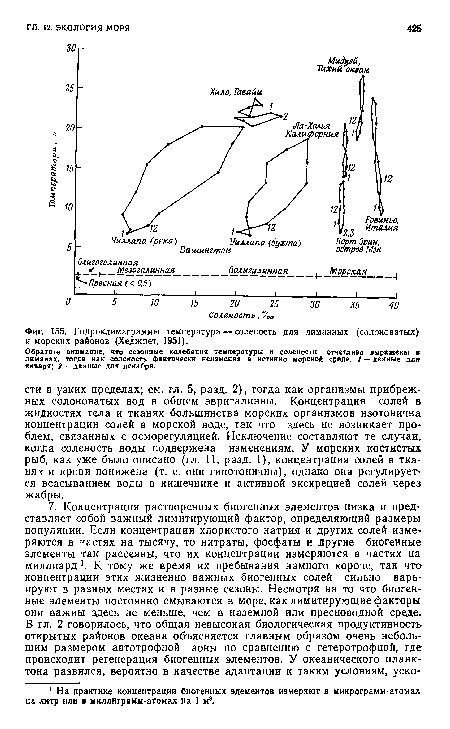 |
Далее
| Теоретическое влияние на фитопланктон шести лимитирующих факторов (вверху). |
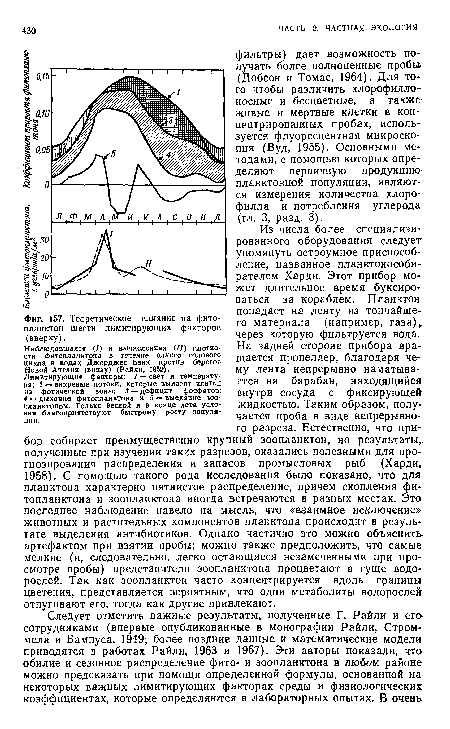 |
Далее
| Морская биота (масштаб не соблюден). |
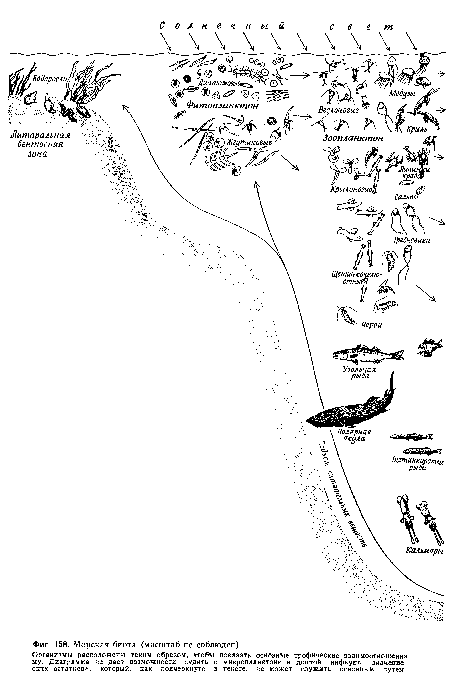 |
Далее
| Фитопланктонные продуценты моря. |
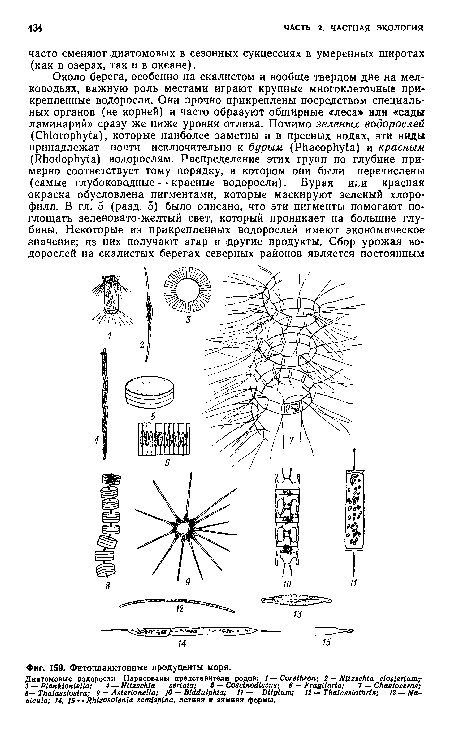 |
Далее
| Фитопланктонные продуценты моря. |
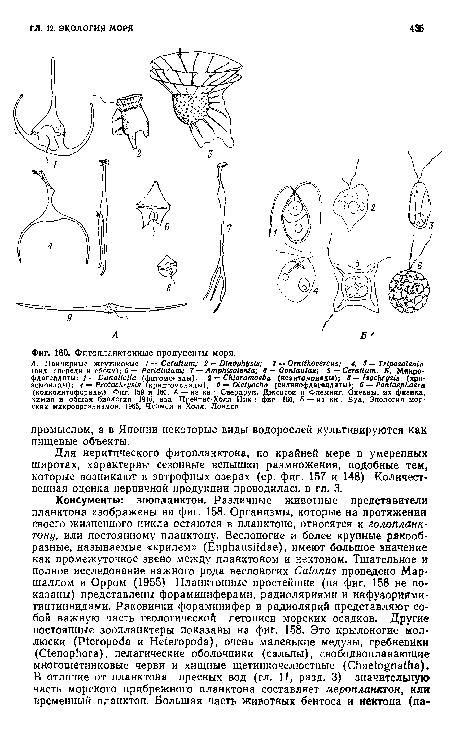 |
Далее
| Личинки, составляющие временный планк тон (меропланктон) морей |
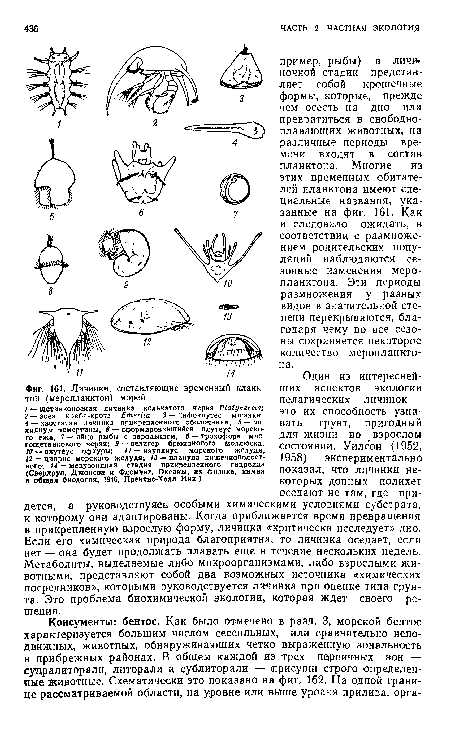 |
Далее
| Схематический разрез песчаного берега (вверху) и скалистого берега (внизу) у Бофорта, Сев. Каролина. |
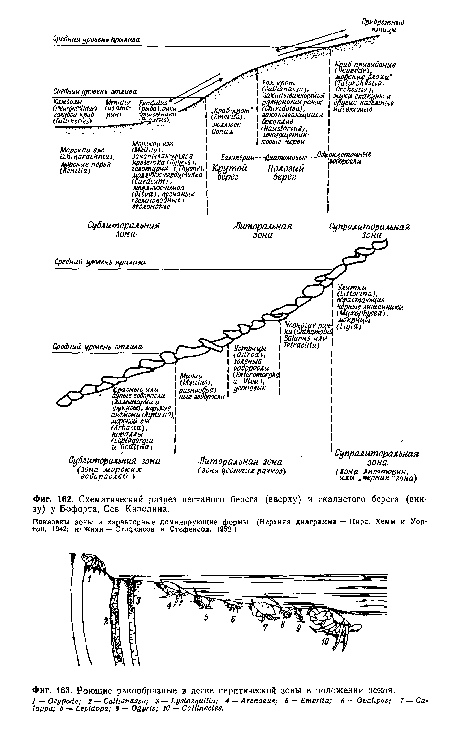 |
Далее
| Роющие ракообразные в песке иеритической зоны в положении покоя. |
|
Далее
| Разрез донных морских осадков, показывающий три зоны, часто различно окрашенные (Фенчель, 1969). |
 |
Далее
| Экосистема мангровых зарослей. |
 |
Далее
| Тайна звукорассеивающего, или глубинного рассеивающего, слоя раскрыта (Бэкус и др., 1968). |
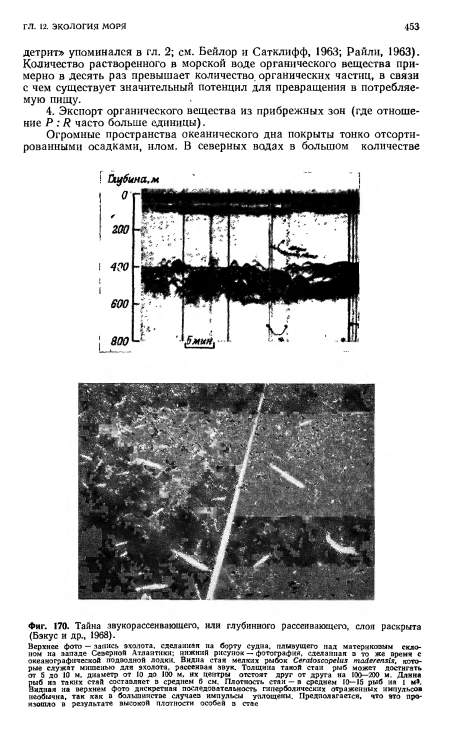 |
Далее
| Схема циркуляции в частично перемешиваемом лимане. |
 |
Далее
| Жизненный цикл креветки, использующей лиман как место откорма молоди. |
 |
Далее
| Зональность в засоленном марше одного из лиманов в Джорджии. |
 |
Далее
| Длинный узкий лист взморника (.Zostera), разрезанный на четыре часги» так что видно развитие эпифитных водорослей на более старых участках (на фиг. справа). |
|
Далее
| Защитная дамба» разрушила наиболее важную часть зоны откорма молоди в лимане, но дала возможность строить здесь жилые здания, подверженные, однако, действию ураганов и штормов (Аллен, 1964; В. Одум, 19706). |
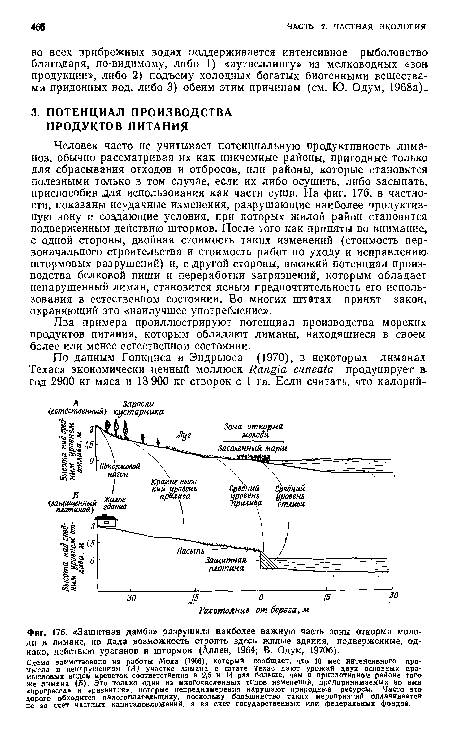 |
Далее
| Биогеографические области земного шара по данным геоботанических и зоогеографических карт. |
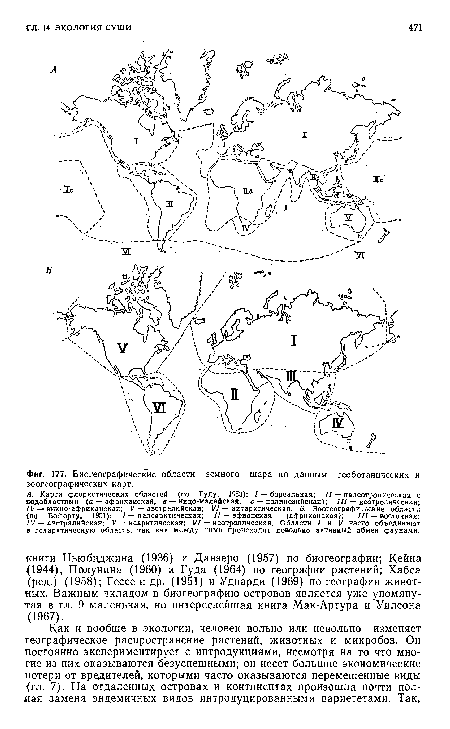 |
Далее
| Жизненные формы наземных растений по Раункиеру. |
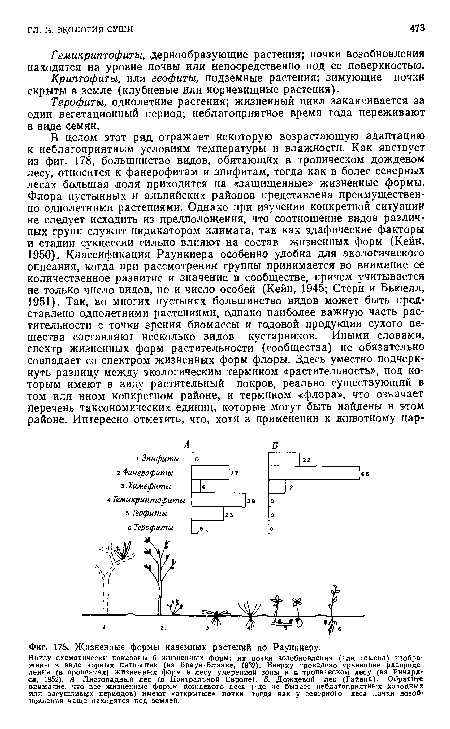 |
Далее
| Живой и неживой компоненты луговой почвы (из Тышлера, 1955). |
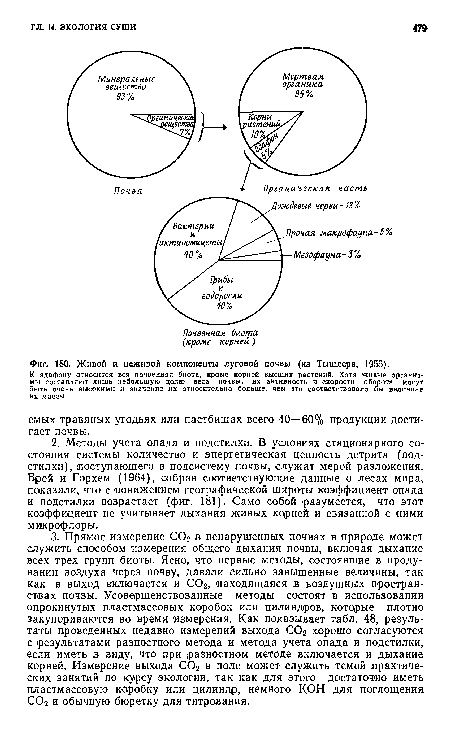 |
Далее
| Годовой опад листьев в лесах, расположен ных на разных широтах |
 |
Далее
| Уменьшение веса подстилки и потеря ею биогенных веществ (по Кроссли и Уиткемпу, 1964). |
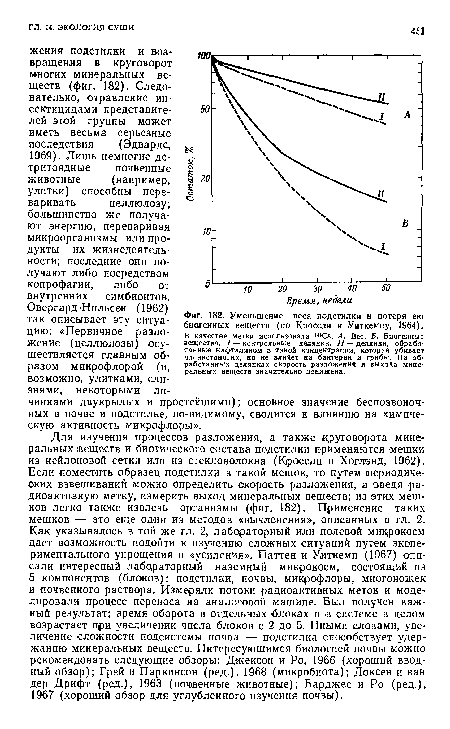 |
Далее
| Главные биомы земного шара. |
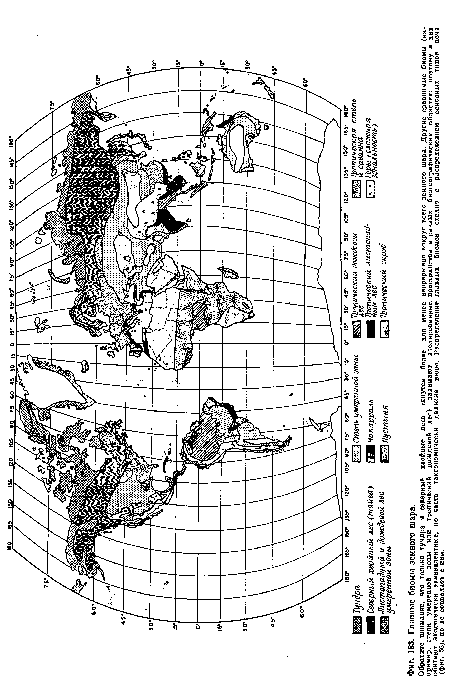 |
Далее
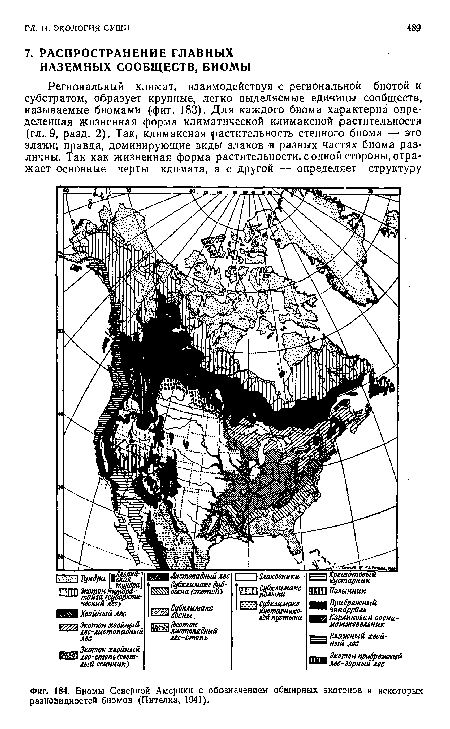
| Биомы Северной Америки с обозначением обширных экотонов и некоторых разновидностей биомов (Пителка, 1941). |
 |
Далее
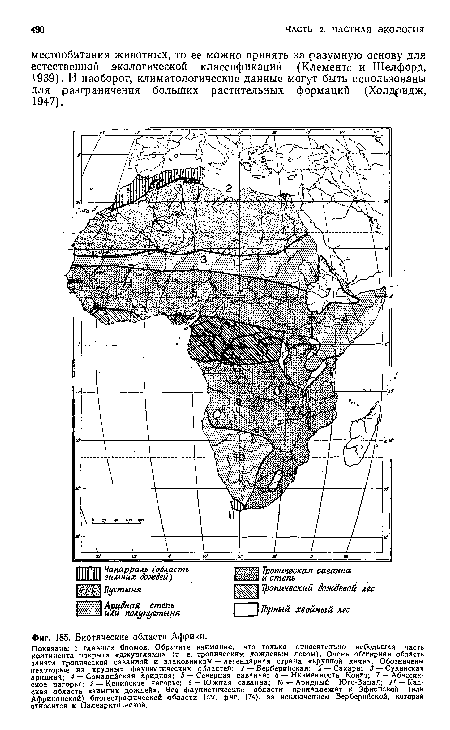
| Биотические области Африки. |
 |
Далее
| Пример влажного хвойного леса, который называют часто дождевым лесом умеренной зоны; заповедник Олимпик (Вашингтон). |
 |
Далее
| Климатический и эдафический климаксы на востоке США. |
 |
Далее
| Остаток пирогенного климаксного леса длиннохвойной сосны близ Томас-вилла (Джорджия). |
 |
Далее
| Естественный злаковник умеренной зоны в центральной части Северной Америки с двумя характерными видами травоядных млекопитающих. |
 |
Далее
| Два климаксных злака биома злаковников в- Северной Америке. |
 |
Далее
| Тропическая саванна в Африке. |
 |
Далее
| Пустыни на западе Северной Америки |
 |
Далее
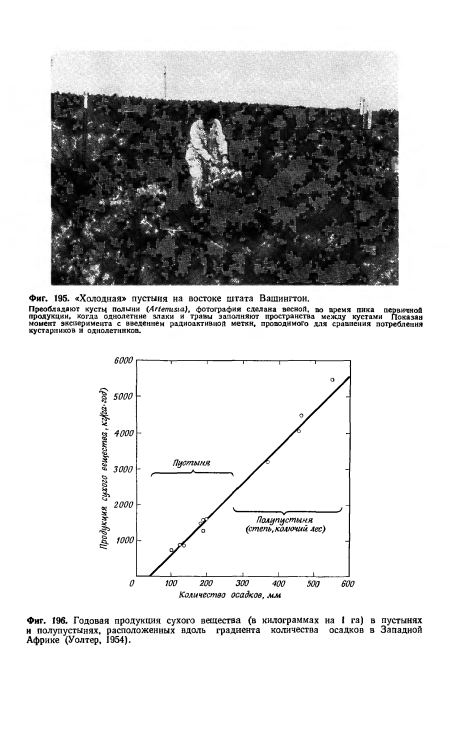
| Годовая продукция сухого вещества (в килограммах на 1 га) в пустынях и полупустынях, расположенных вдоль градиента количества осадков в Западной Африке (Уолтер, 1954). |
 |
Далее
| Биом карликовой сосны и можжевельника в Аризоне. |
 |
Далее
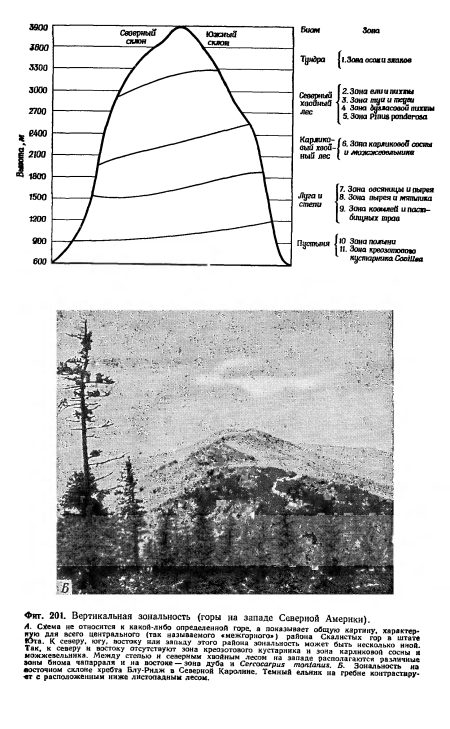
| Вертикальная зональность (горы на западе Северной Америки). |
 |
Далее
| Альтернативные схемы истощения запасов полезных ископаемых. |
 |
Далее
| Зависимость урожая пищевых культур от применения удобрений (/), пестицидов (//) и затраты мощностей (///) при выращивании и сборе урожая. |
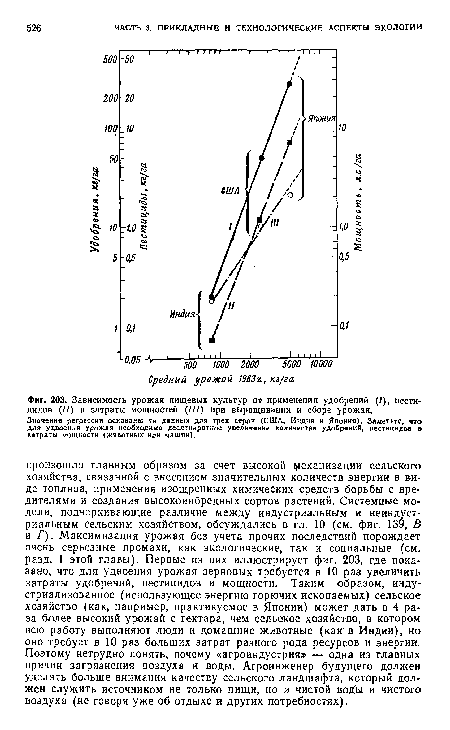 |
Далее
| Саженый сосновый лес. |
 |
Далее
| Естественный сосновый бор. |
 |
Далее
| Классификация земельных угодий (объяснения в тексте). |
 |
Далее
| План развития городского района с населением в 110 000 чел. на плошади 18 000 га, обеспечивающий сохранение качества окружающей среды и красот природы и предусматривающий достаточно места для отдыха и удаления загрязнений. |
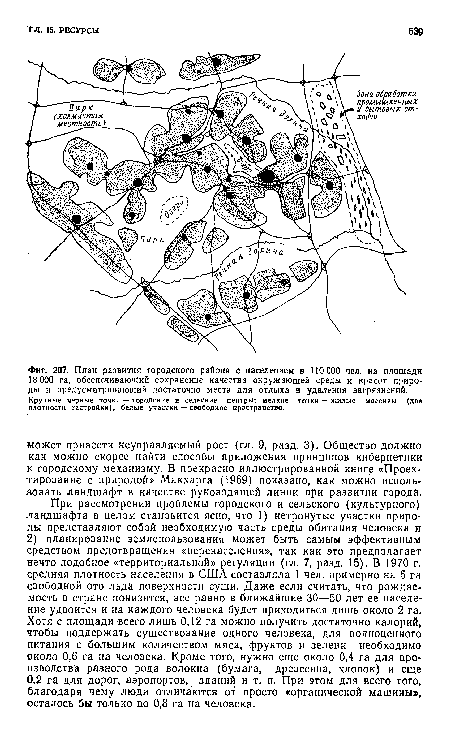 |
Далее
| Цена загрязнений. |
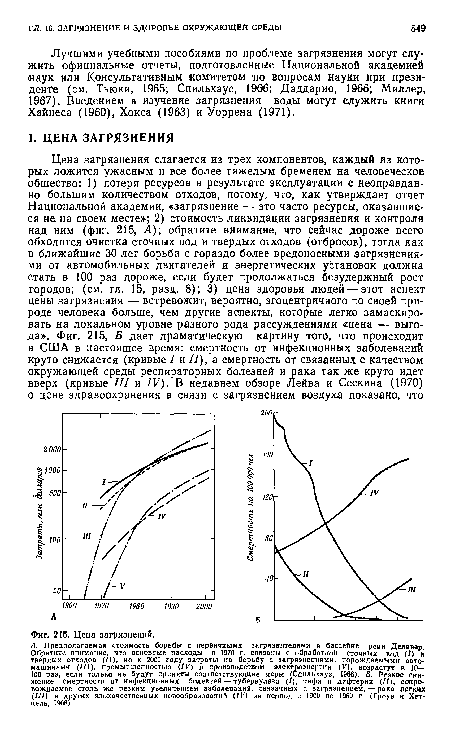 |
Далее
| Влияние на экосистему загрязнений двух типов—нестойких органических (/) и стойких токсичных (II). |
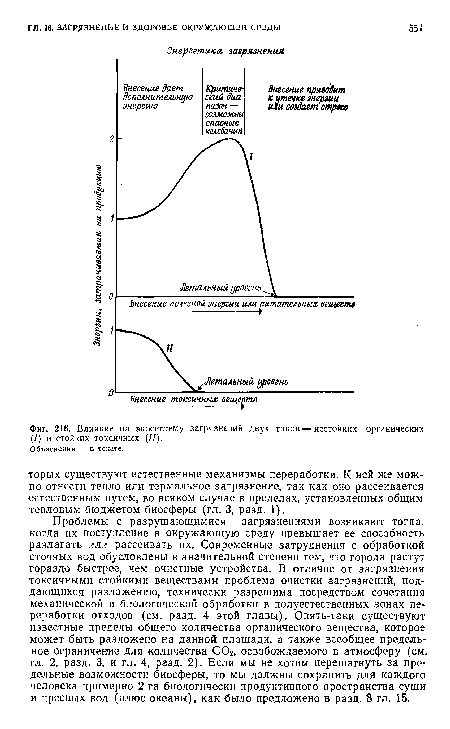 |
Далее
| Три стадии обработки сточных вод и других органических отходов. |
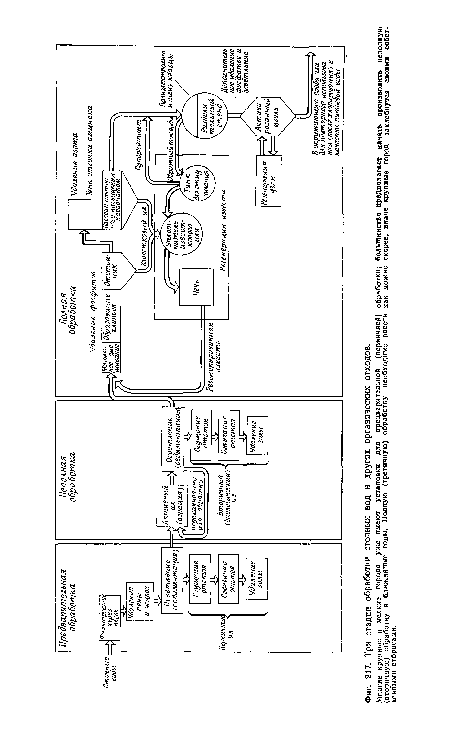 |
Далее
| Обработка отходов от нефтеочистительного завода в Оклахоме довольно дешевым способом (Копленд и Доррис, 1964). |
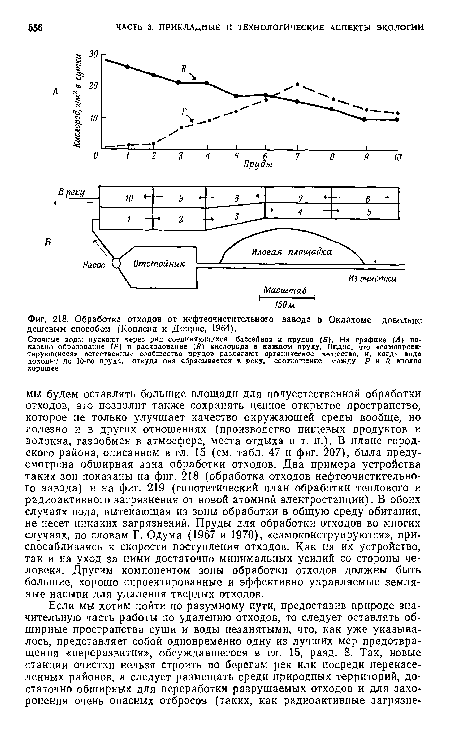 |
Далее
| Схема зоны обработки отходов для будущей атомной электростанции (а), расположенной в естественном водосборном бассейне (очерчен пунктиром). |
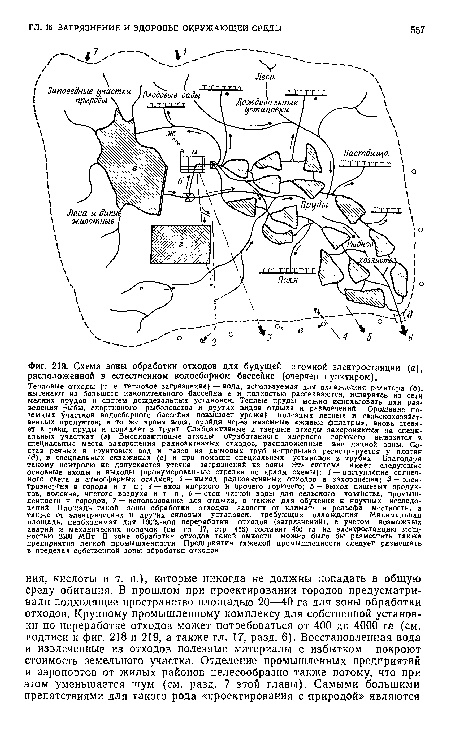 |
Далее
| Сравнение трех типов ионизирующего излучения, представляющих наибольший экологический интерес. |
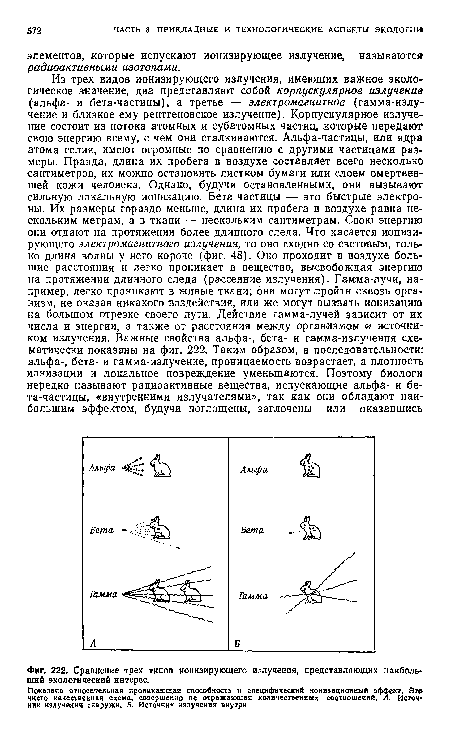 |
Далее
| Сравнительная чувствительность трех разных организмов к единичной острой дозе рентгеновских или гамма-лучей (объяснения — в тексте). |
 |
Далее
| Влияние на дубово-сосновый лес градиента гамма-облучения от высокоактивного неподвижного источника. |
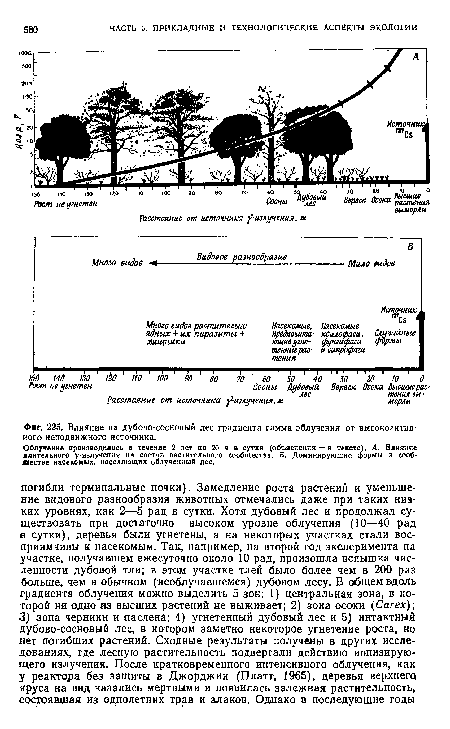 |
Далее
| Накопление стронция-90 в разных частях сетн питания одного небольшого канадского озера, получающего низкоактивные отходы (Офель, 1963). |
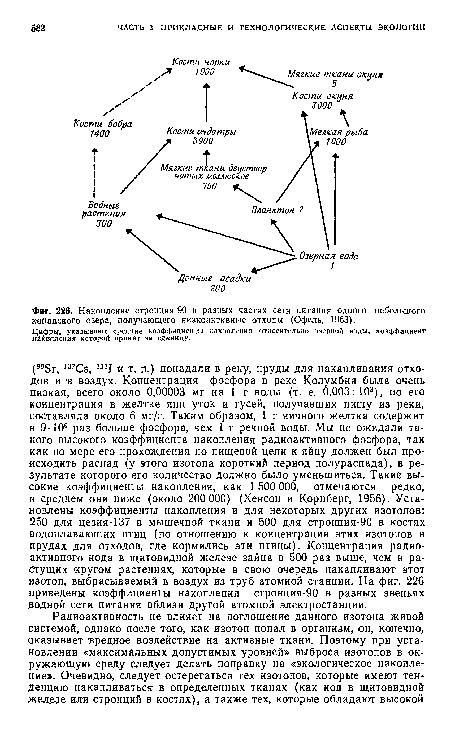 |
Далее
| Применение радиоактивных меток для составления схем пищевых цепей в интактных природных сообществах. |
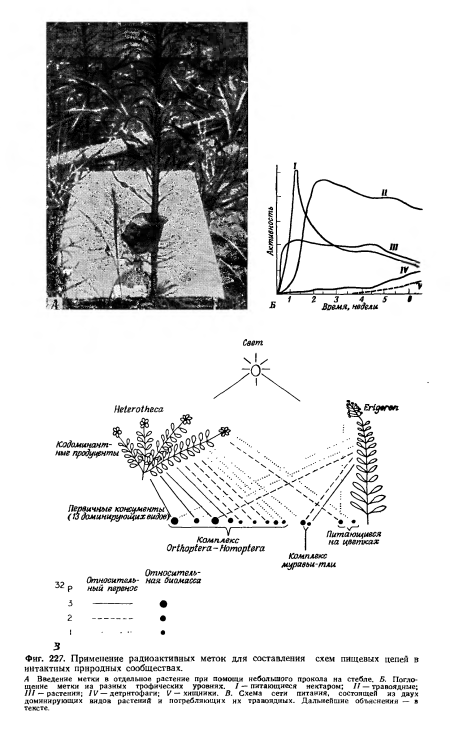 |
Далее
| Захоронение в грунт высокоактивных жидких отходов на Ханфордском заводе (Браун, Паркер и Смит, 1956) |
 |
Далее
| При интерпретации фотографии (И. Ф.) по аналогии с теоретико-информа-ционной моделью энтропия сложной структуры уменьшается по мере извлечения информации. |
 |
Далее
| Установка для измерения оптической плотности, регистрирующая ослабление проходящего света. |
 |
Далее
| Картк части угодий сельскохозяйственной фермы университета Пардью (конец июня 1966 г.). |
 |
Далее
| Расшифровка карты посевов пшеницы. |
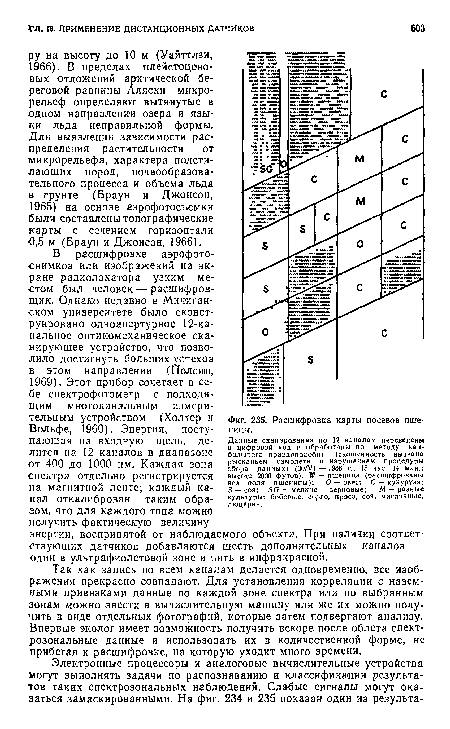 |
Далее
| Карта растительности и аэрофотосъемка структуры растительности в Чант-хабури (Таиланд) |
 |
Далее
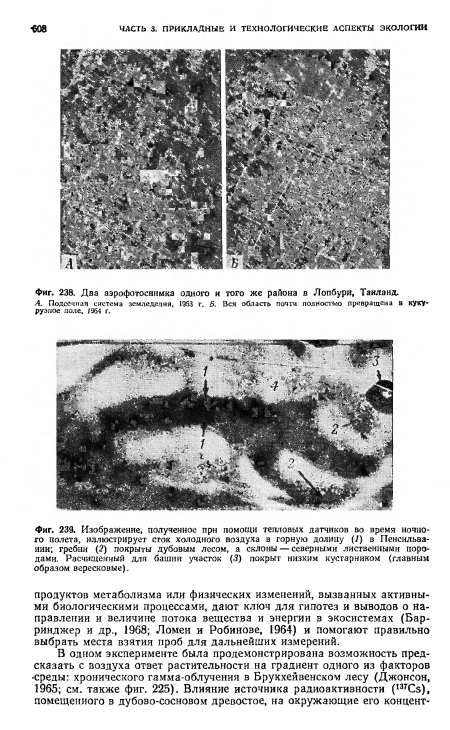
| Два аэрофотоснимка одного и того же района в Лопбури, Таиланд. |
 |
Далее
| Изображение, полученное прн помощи тепловых датчиков во время ночного полета, иллюстрирует сток холодного воздуха в горную долину (1) в Пенсильвании; гребни (2) покрыты дубовым лесом, а склоны — северными лиственными породами. Расчищенный для башни участок (3) покрыт низким кустарником (главным образом вересковые). |
|
Далее
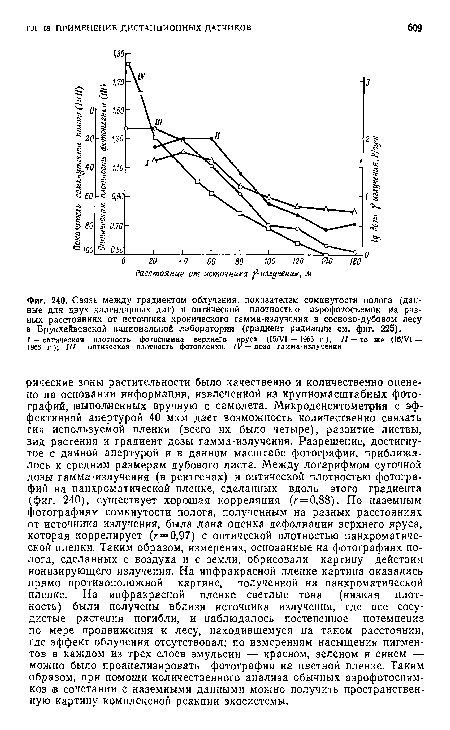
| Связь между градиентом облучения, показателем сомкнутости полога (данные для двух календарных дат) и оптической плотностью аэрофотосъемок иа разных расстояниях от источника хронического гамма-излучения в сосново-дубовом лесу в Брукхейвеиской национальной лаборатории (градиент радиации см. фиг. 225). |
 |
Далее
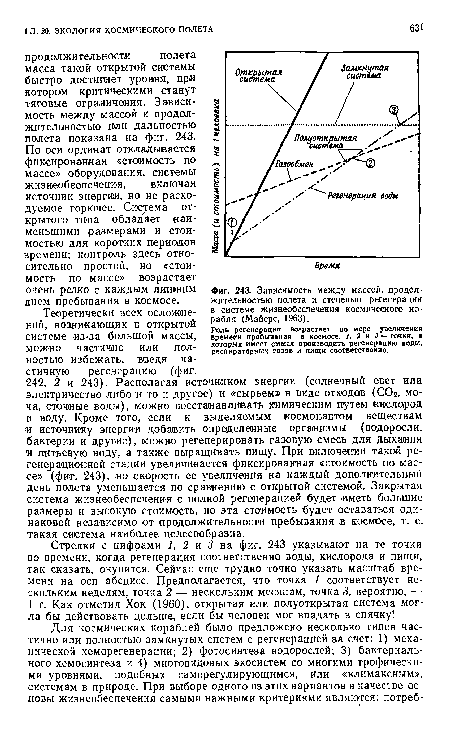
| Зависимость между массой, продолжительностью полета и степенью регенерации в системе жизнеобеспечения космического корабля (Майерс, 1963). |
 |
Далее
| Макет полной системы жизнеобеспечения, в которой механическими и Химическими способами производится регенерация воды и газов |
 |
Далее
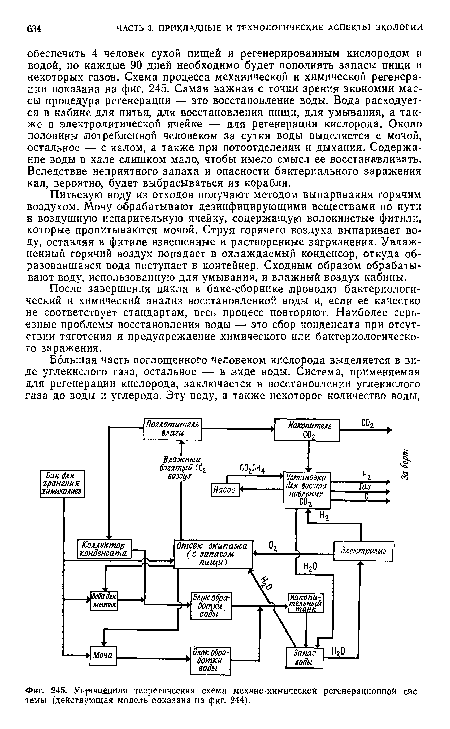
| Упрещеняая теоретическая схема механо-химической регенерационной системы (действующая модель показана на фиг. 244). |
 |
Далее
| Упрощенная схема биорегенерационной системы, основанной либо на хемосинтезирующих бактериях, либо на фотосинтезирующих растениях. |
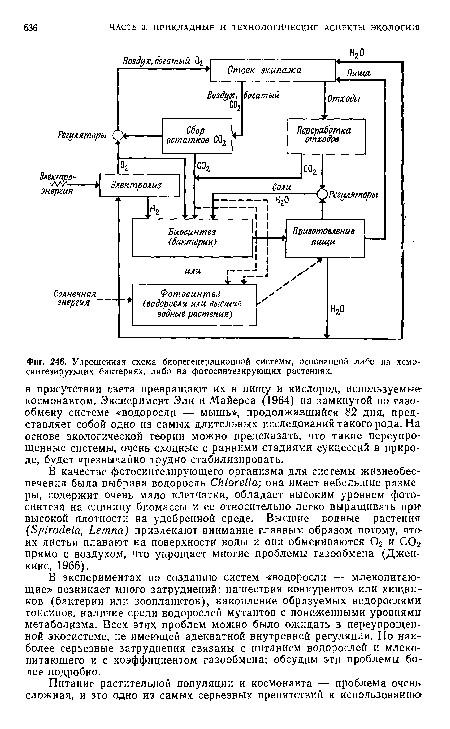 |
Далее
| Два основных «варианта выбора» для будущего роста населения. |
 |
Далее