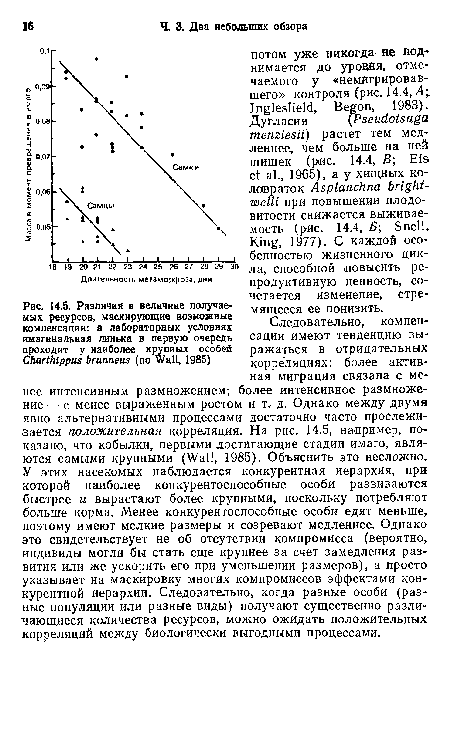
| Различия в величине получаемых ресурсов, маскирующие возможные компенсации |
 |
Далее
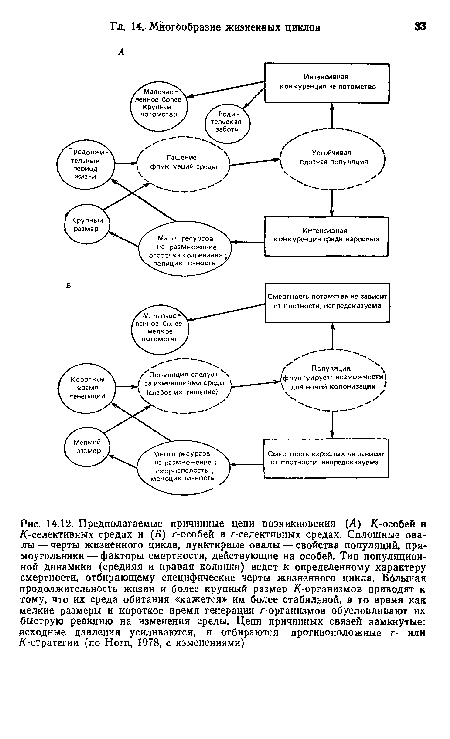
| Предполагаемые причинные цепи возникновения (Л) К-особей в /(-селективных средах и (Б) r-особей в r-селективных средах. Сплошные овалы— черты жизненного цикла, пунктирные овалы — свойства популяций, прямоугольники— факторы смертности, действующие на особей. Тип популяционной динамики (средняя и правая колонки) ведет к определенному характеру смертности, отбирающему специфические черты жизненного цикла. Большая продолжительность жизии и более крупный размер /(-организмов приводят к тому, что их среда обитания «кажется» им более стабильной, в то время как мелкие размеры и короткое время генерации r-организмов обусловливают их быструю реакцию на изменения среды. Цепи причинных связей замкнутые |
 |
Далее
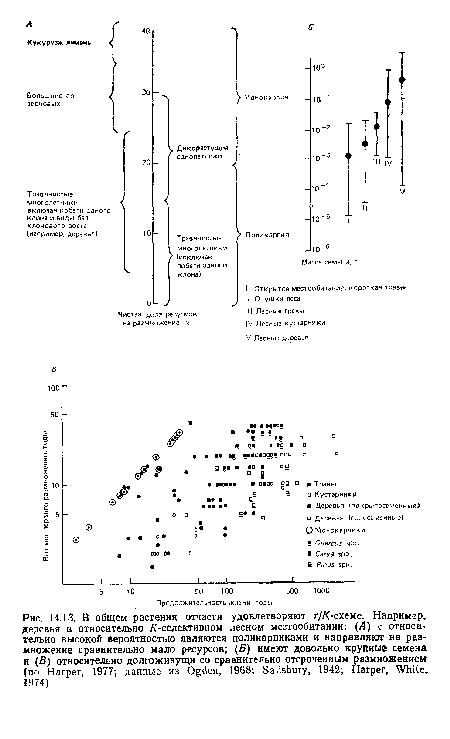
| В общем растения отчасти удовлетворяют г ¡К-схеме. Например, деревья в относительно /С-селектнвном лесном местообитании |
 |
Далее
| Зависимость между остаточной репродуктивной ценностью и размером литторин на валунах и в ¿целях. Типы местообитаний характеризуются градиентом этой зависимости, как показано в тексте (размножение в обоих случаях начинается при одном и том же градиенте). Показаны черты жизненных циклов, выгодные в том или ином местообитании и соответствующие полевым наблюдениям (табл. 14.3) |
 |
Далее
| Аллометрическая зависимость между высотой дерева и диаметром ствола на высоте 1,525 м для 576 отдельных деревьев, представляющих почти, все виды, произрастающие в Америке (по McMahon, 1973) |
 |
Далее
| Изменения размера популяции лютика ползучего на участке С (см. табл. 15.1). О — суммарное пополнение за счет прорастающих семян и клонального роста; # — суммарные потери; □ — результирующая динамика популяции (по Sarukhan, Harper, 1973) |
 |
Далее
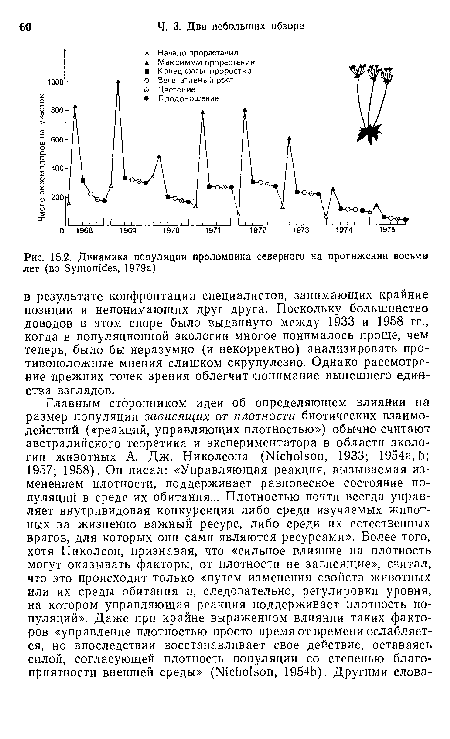
| Динамика популяции проломника северного на протяжении восьми лет (по Symonides, 1979а) |
 |
Далее
| Регулирование численности популяции (/1) не зависящей от плотно -сти рождаемостью и зависящей от плотности смертностью; (£) зависящей от плотности рождаемостью и не зависящей от плотности смертностью; (В) зависящими от плотности рождаемостью и смертностью. Размер популяции растет, когда рождаемость выше смертности, и падает, когда смертность выше рождаемости. N° — устойчивый равновесный размер популяции, определяемый как интенсивностью не зависящих от плотности процессов, так и интенсивностью и углом наклона графика зависящих от плотности процессов |
 |
Далее
| Временные изменения разных й-факторов в канадских популяциях колорадского жука (данные Нагсоиг!, 1971) |
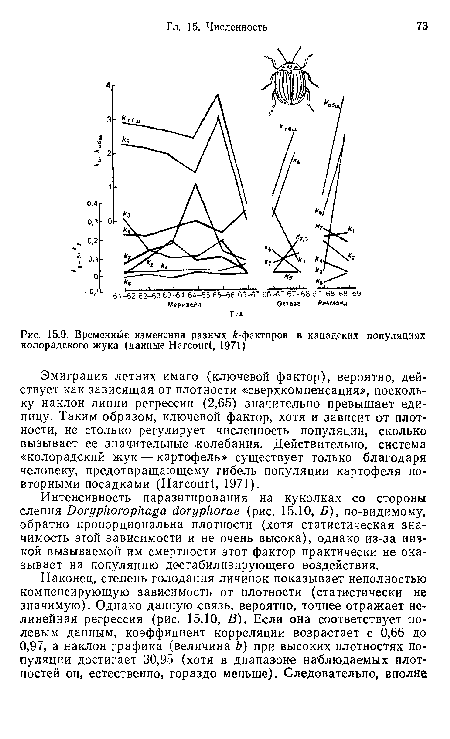 |
Далее
| А. Зависящая от плотности эмиграция «летних» имаго колорадского жука (наклон 2,65). Б. Обратно пропорциональная зависимость от плотности паразитизма на куколках (наклон —0,11). В. Зависимость от плотности в случае личиночного голодания (наклон прямой 0,37; наклон в конце кривой 30,95) (данные Нагсош!, 1971) |
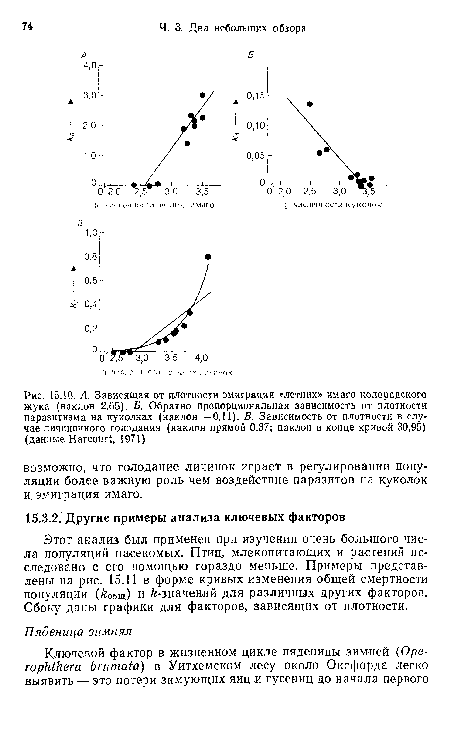 |
Далее
| Возрастная структура (А) и кривые выживания (Б) для популяций отдельных листьев лютика ползучего (Peters, 1980) |
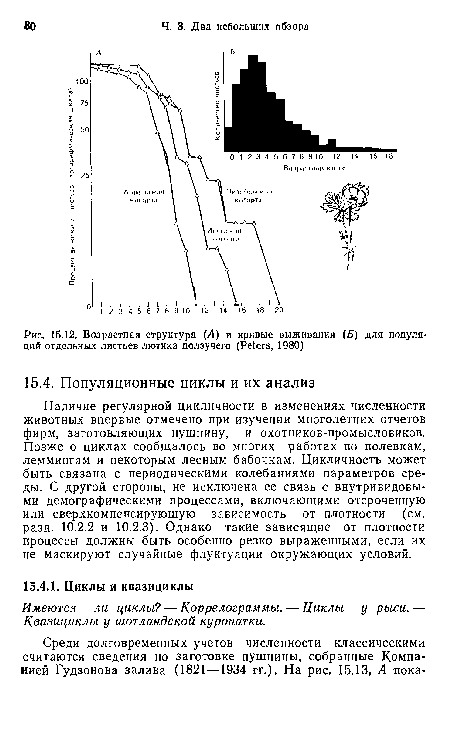 |
Далее
| Оценки плотности леммингов в береговой тундре окрестностей Барроу (Аляска) для периода 20 лет (по ЪаЫ] е£ а1., 1980) |
![Оценки плотности леммингов в береговой тундре окрестностей Барроу (Аляска) для периода 20 лет (по ЪаЫ] е£ а1., 1980)](/static/pngsmall/228045074.png) |
Далее
| Схема событий по гипотезе восстановления питательных веществ (по Schultz, 1964, 1969) |
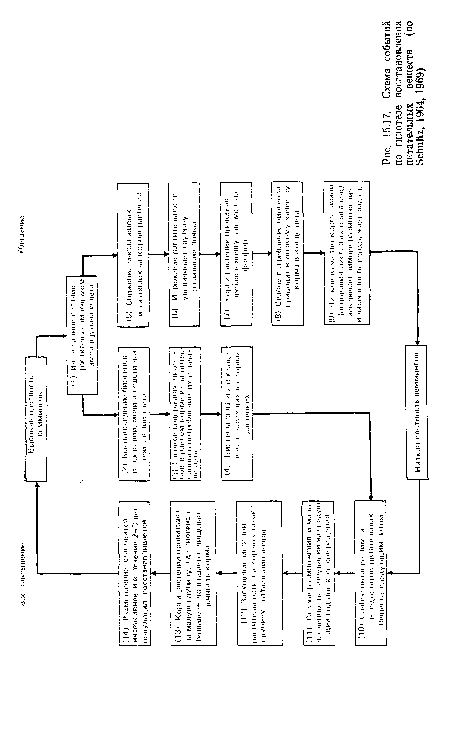 |
Далее
| Оценки плотности хищников на протяжении стандартного цикла леммингов в окрестностях Барроу. Периоды таяния снега и наступления холодов указаны промежутками между летом и зимой (по Ва№1 е1 а!., 1980) |
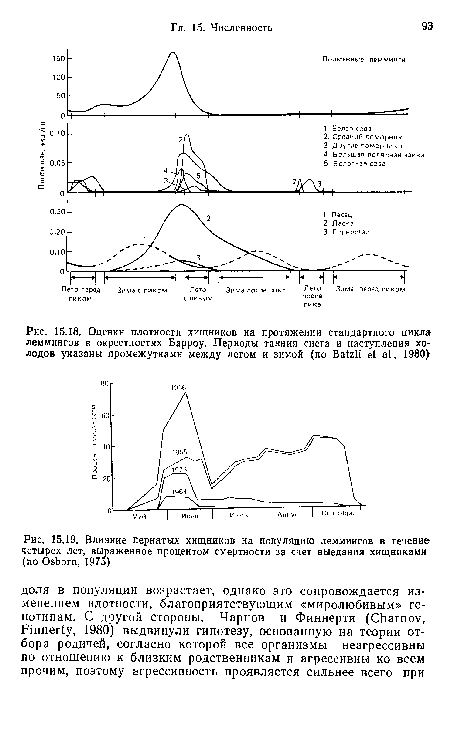 |
Далее
| Влияние пернатых хищников на популяцию леммингов в течение четырех лет, выраженное процентом смертности за счет выедания хищниками (по Osborn, 1975) |
|
Далее
| Последствия удаления разных видов на пяти участках Кор-Банкс. На каждом рисунке площадь центрального (жирно очерченного) круга соответствует относительному обилию (проценту покрытия) вида до его удаления. Площади внешних кругов — обилие видов в контроле. Площади кругов, вдающихся внутрь центрального, —повышение обилия видов, оставшихся после удаления |
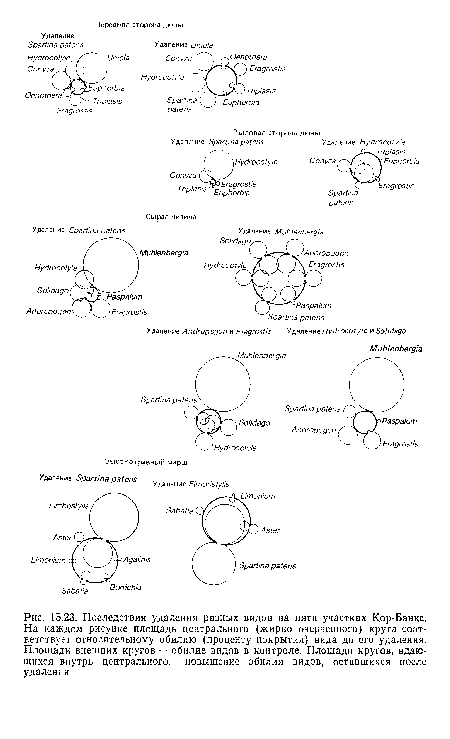 |
Далее
| Озеро Мун-Дарра (север шт. Квинсленд, Австралия). Вверху |
 |
Далее
| Борьба с опунцией с помощью кактусовой огневки. Математическая модель (Caughley, Lawton, 1981) точно описывает падение обилия кактуса за два года с 5000 экз./акр (1 акр — 0,4 га) перед выпуском бабочек до стабильного значения И экз./акр. Верхняя кривая — обилие опунции, нижняя — численность бабочек после интродукции в «нулевой» год |
 |
Далее
| Можно обнаружить иерархию местообитаний, как бы вставленных друг в друга |
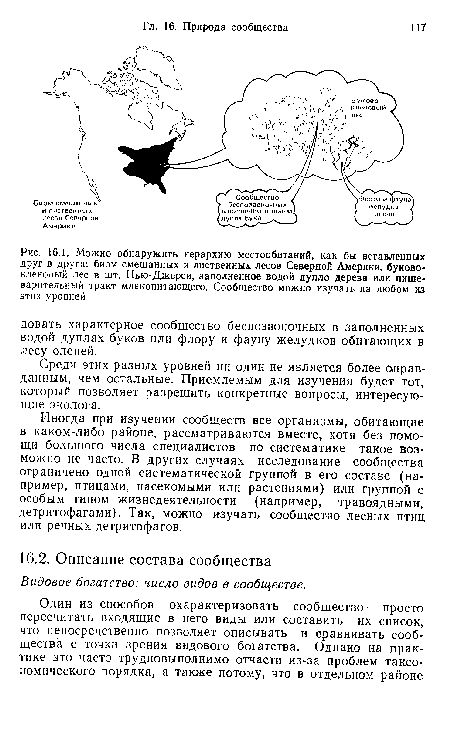 |
Далее
| Зависимость между видовым богатством и численностью отдельных организмов в двух гипотетических сообществах. В сообществе А видовое богатство гораздо выше, чем в сообществе Б |
 |
Далее
| Примеры рангового распределения обилий для трех гипотетических сообществ. Указаны соответствующие каждой кривой значения равномерности распределения (Е, по Симпсону) |
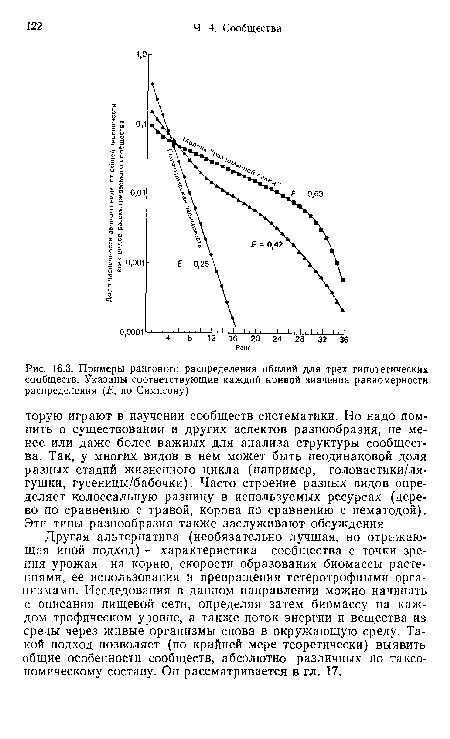 |
Далее
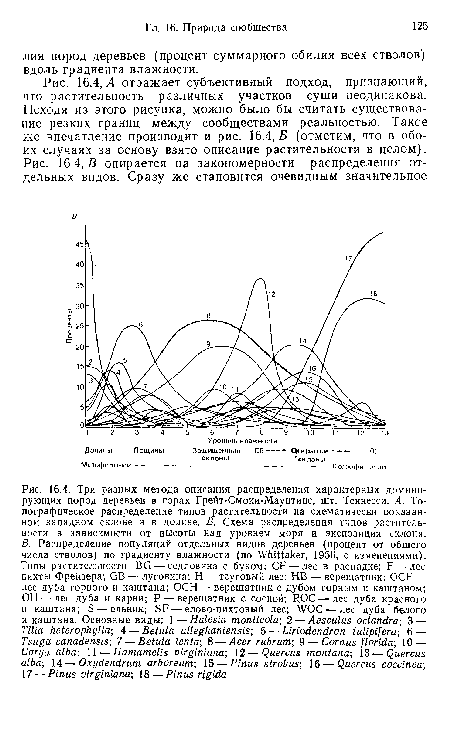
| Л показывает характерные ассоциации на горном склоне так, как будто между ними проходят четкие границы. На склоне условия для роста растений неоднородны, причем на распределение пород деревьев, по-видимому, особенно сильно влияют высота над уровнем моря и влажность. На рис. 16.4, Б приведены доминирующие ассоциации при различных значениях этих факторов. Наконец, на рис. 16.4,5 представлено распределение оби- |
 |
Далее
| Л отражает субъективный подход, признающий, что растительность различных участков суши неодинакова. Исходя из этого рисунка, можно было бы считать существование резких границ между сообществами реальностью. Такое же впечатление производит и рис. 16.4,5 (отметим, что в обоих случаях за основу взято описание растительности в целом). |
 |
Далее
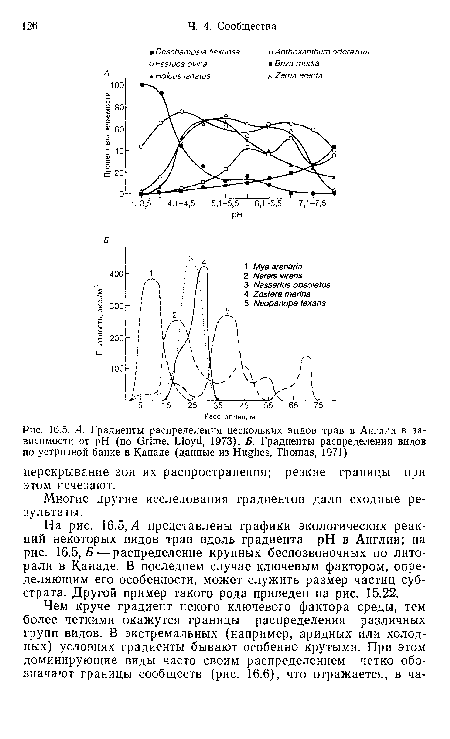
| А. Градиенты распределения нескольких видов трав в Англии в зависимости от pH (по Grime, Lloyd, 1973). Б. Градиенты распределения видов по устричной банке в Канаде (данные из Hughes, Thomas, 1971) |
 |
Далее
| Резко или нет происходит смена одного предсказуемого сообщества другим, зависит от пространственного распределения важнейших для организмов условий среды. А. Если условия меняются ступенчато, группы видов могут быть резко отграничены одна от другой. Б. Если условия изменяются постепенно, относительные обилия доминирующих видов меняются так же |
 |
Далее
| Состав сообществ (каждое из 5-6 видов) в четырех местах, располагающихся вдоль средового градиента. По оси Y откладывается обилие каждого из видов (А—Н), выражаемое процентом общей численности или биомассы, а по оси X — числовые значения какого-либо фактора, например высота над уровнем моря, влажность почвы, размер частиц почвы и т. д. |
 |
Далее
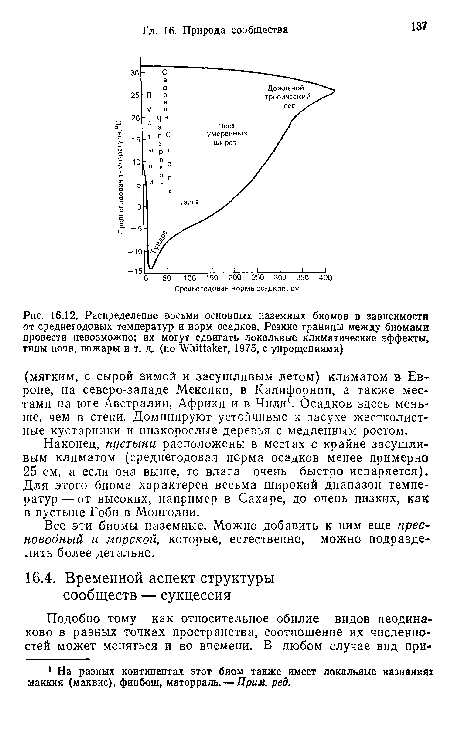
| Распределение восьми основных наземных биомов в зависимости от среднегодовых температур и иорм осадков. Резкие границы между биомами провести невозможно; их могут сдвигать локальные климатические эффекты, типы почв, пожары и т. д. (по Whittaker, 1975, с упрощениями) |
 |
Далее
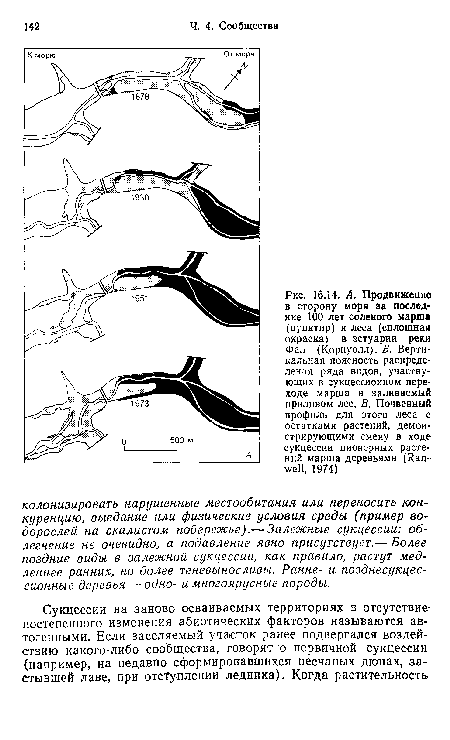
| А. Продвижение в сторону моря за последние 100 лет соленого марша (пунктир) и леса (сплошная окраска) в эстуарии реки Фал (Корнуолл). Б. Вертикальная поясность распределения ряда видов, участвующих в сукцессиоином переходе марша в заливаемый приливом лес. В. Почвенный профиль для этого леса с остатками растений, демонстрирующими смеиу в ходе сукцессии пионерных растений марша деревьями (Ran-well, 1974) |
 |
Далее
| Сукцессия растительности после отступления ледника в районе бухты Глейшер (Аляска) и изменения общего содержания азота в почвах (но Crocker, Major, 1955) |
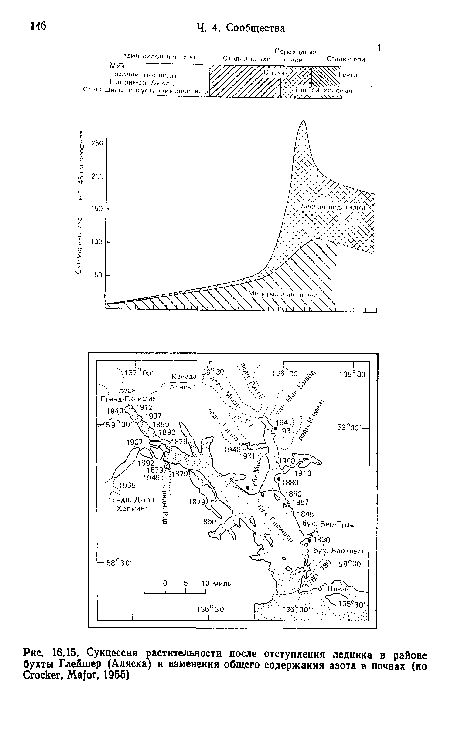 |
Далее
| Средний процент (± стандартная ошибка) покрытия бетонных блоков, установленных на литорали в сентябре 1974 г., каждым из пяти видов водорослей (Sousa, 1979а) |
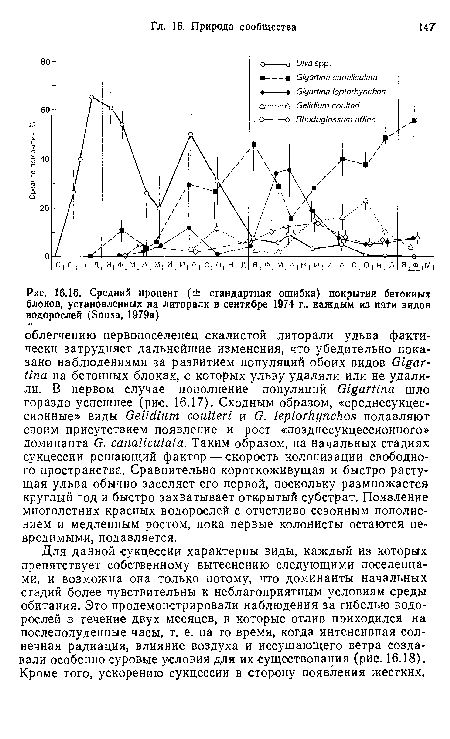 |
Далее
| Кривые выживания для пяти видов водорослей в течение двух месяцев начиная с 5 декабря 1976 г., когда послеполуденные отливы создавали суровые условия существования. Изначально было помечено по тридцать экземпляров каждого вида (Sousa, 1979а) |
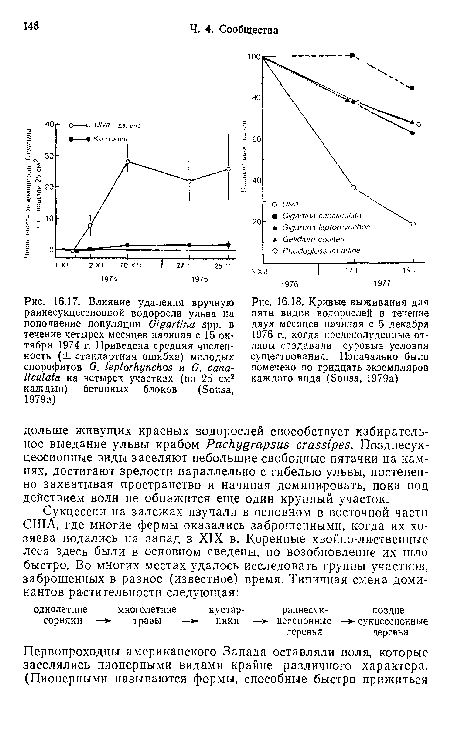 |
Далее
| Три модели механизмов сукцессий (по Connell, Slatyer, 1977, с изменениями) |
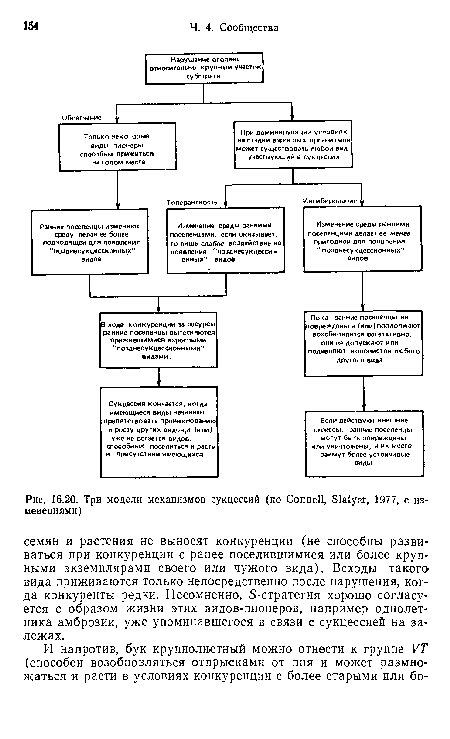 |
Далее
| Широтное распределение чистой первичной продукции для (Л) степных и тундровых экосистем (Cooper, 1975); (Б) зерновых культур (Cooper, 1975); (В) озер (Brylinsky, Mann, 1973) |
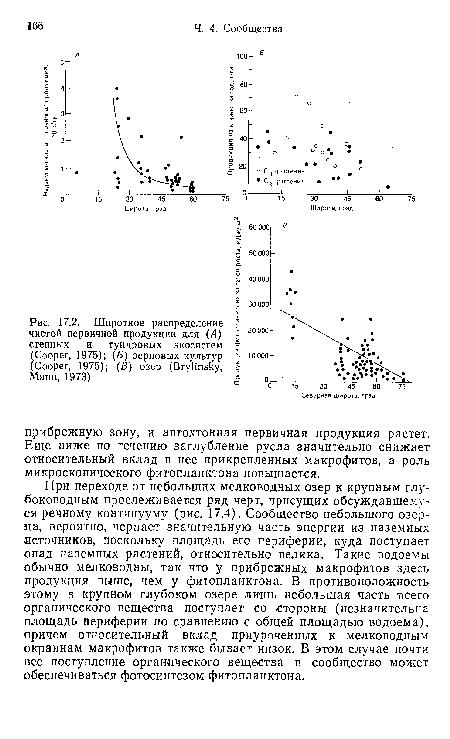 |
Далее
| Изменение энергетической базы сообществ по течению реки |
 |
Далее
| Изменение доли поступления органического вещества с суши, а также доли первичной продукции литорали и планктона в различных водных сообществах |
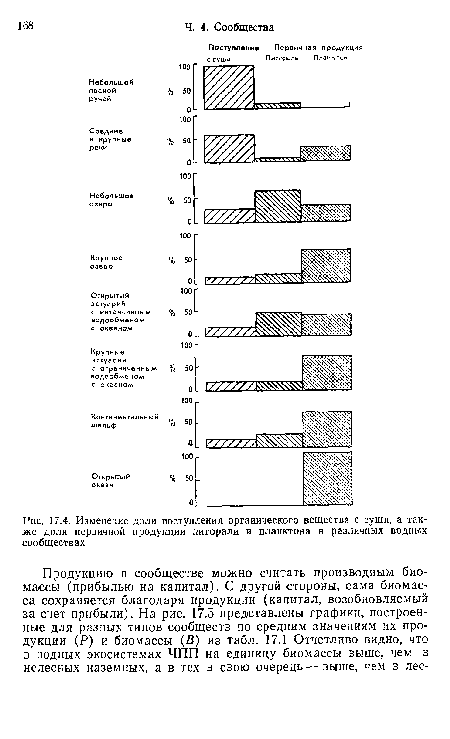 |
Далее
| Схемы реакций растительности на температуру. А. Обобщенный график скорости роста в зависимости от температуры с тремя критическими участками |
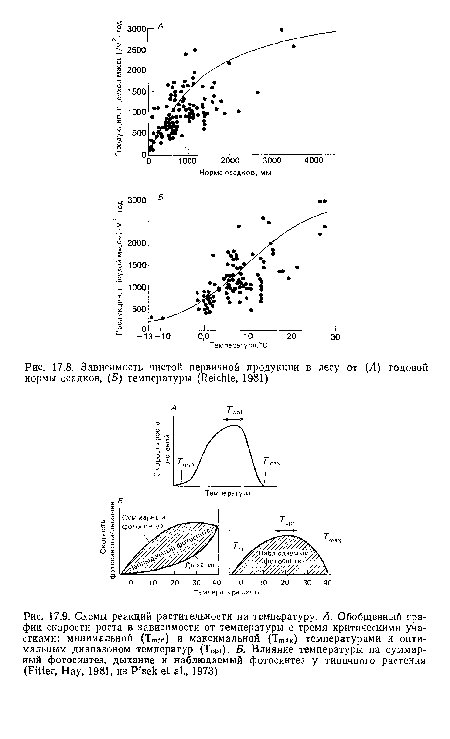 |
Далее
| Зависимость между чистой первичной продукцией листопадных лесов Северной Америки и длительностью вегетационного периода (Leith, 1975) |
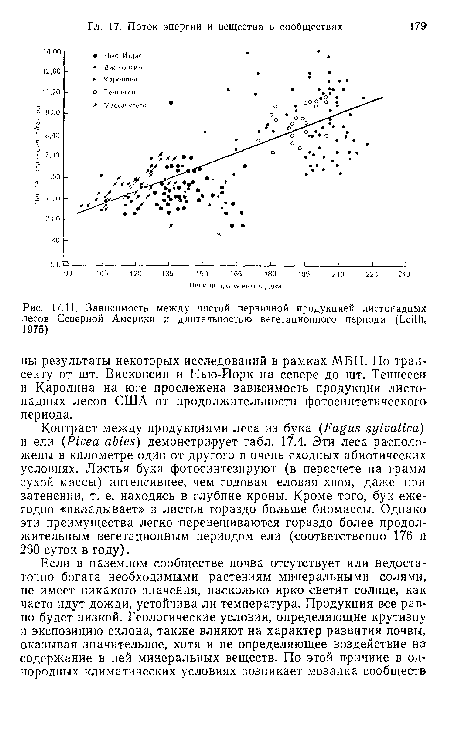 |
Далее
| Зависимость между чистой первичной продукцией фитопланктона в озерах всего земного шара и устойчивой концентрацией в воде фосфора (Schindler, 1978) |
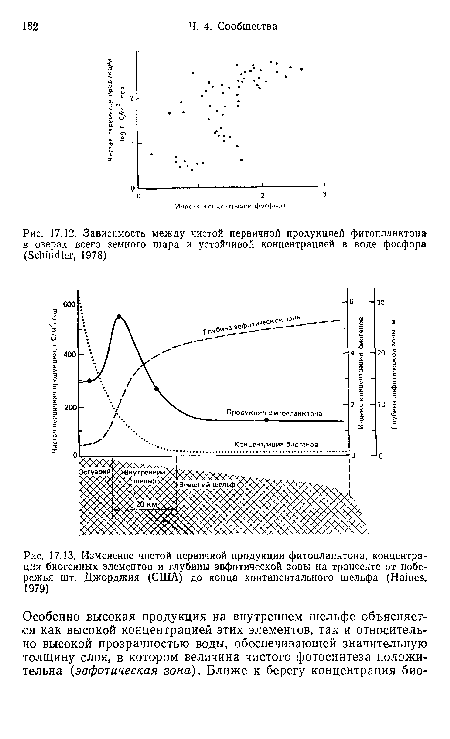 |
Далее
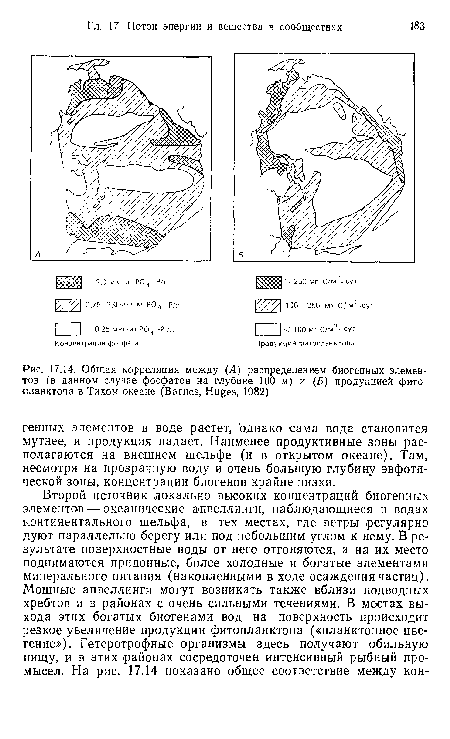
| Изменение чистой первичной продукции фитопланктона, концентрации биогенных элементов и глубины эвфотической зоны на трансекте от побережья шт. Джорджия (США) до конца континентального шельфа (Haines, 1979) |
|
Далее
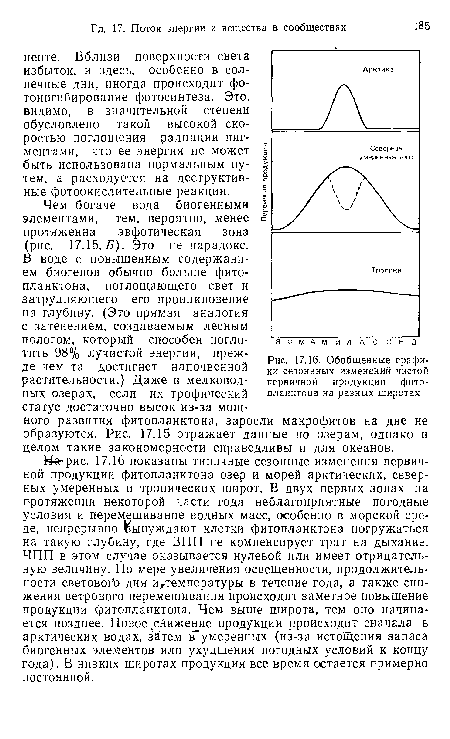
| Общая корреляция между (А) распределением биогенных элементов (в данном случае фосфатов на глубине 100 м) и (Б) продукцией фитопланктона в Тихом океане (Barnes, Huges, 1982) |
 |
Далее
| Обобщенные графики сезоннных изменений чистой первичной продукции фитопланктона на разных широтах |
 |
Далее
| Схема потока энергии в сообществе |
 |
Далее
| Зависимость продукции от интенсивности дыхания для семи групп животных |
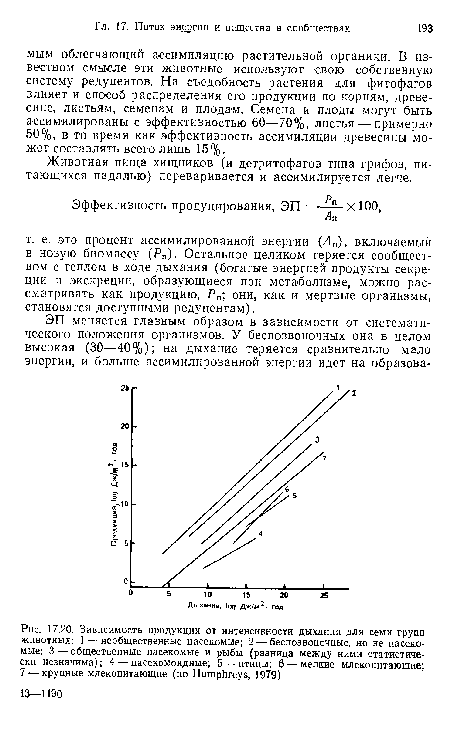 |
Далее
| Схема потока энергии через каждый трофический блок рис. 17.19 и значения эффективностей ассимиляции и продуцирования, использованные Хилом и МакЛином (Heal, MacLean, 1975) в модели степной экосистемы |
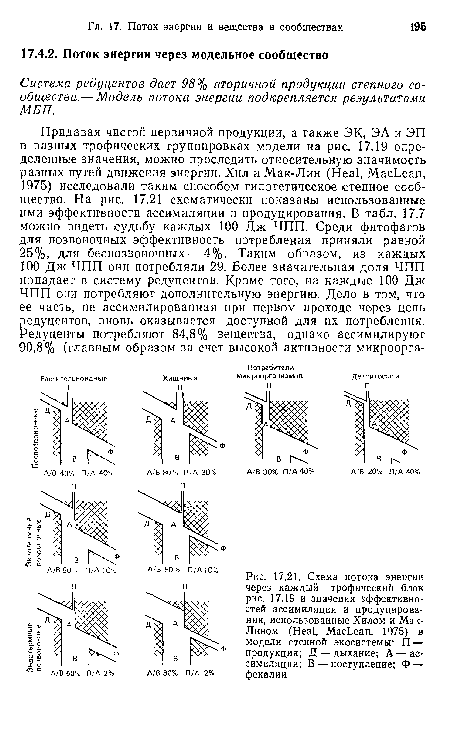 |
Далее
| Зависимость между прогнозируемой и наблюдаемой продукциями гетеротрофов в различных местообитаниях |
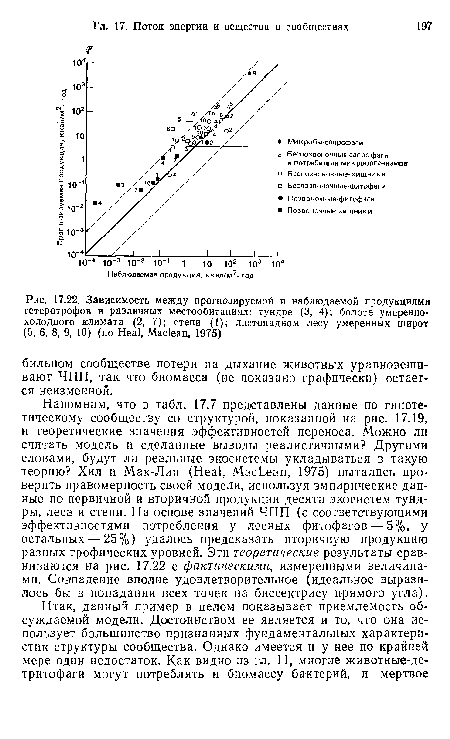 |
Далее
| Общие схемы потоков энергии для |
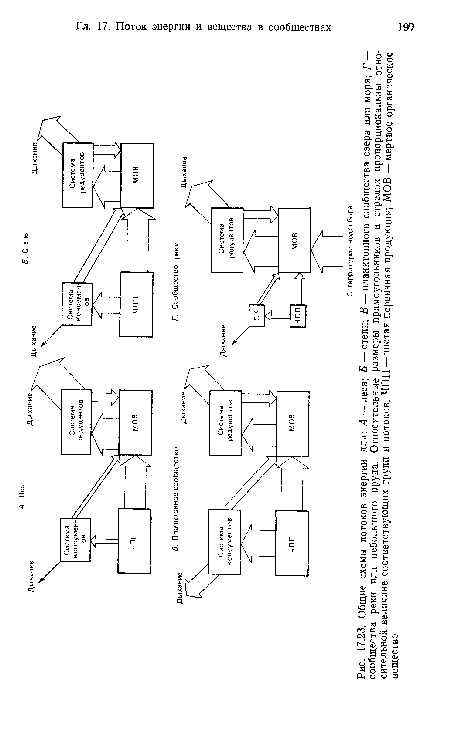 |
Далее
| Диаграмма зависимости между потоком энергии (1) и круговоротом биогенных элементов; 2 — поток биогенных элементов, связанных с органическим веществом; 3 — их свободная неорганическая форма |
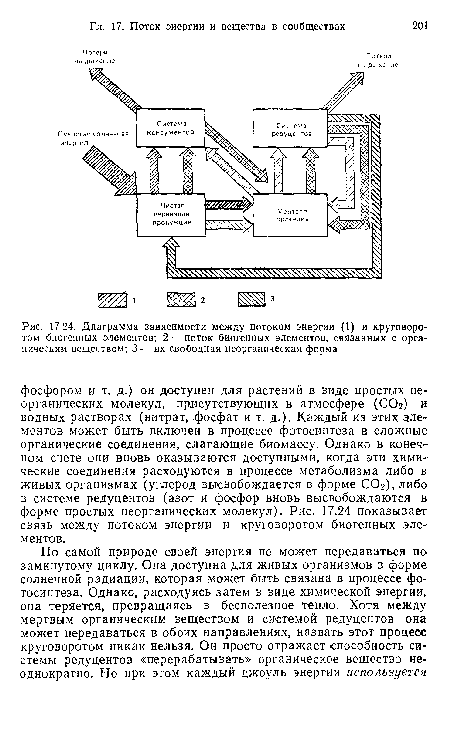 |
Далее
| Годовой баланс азота для ненарушенного леса Хаббард-Брук. Цифры в прямоугольниках соответствуют запасам азота (в килограммах на гектар). Скорости прироста (в скобках) и переноса даны в килограммах азота на гектар за год (Вогтапп е! а!., 1977) |
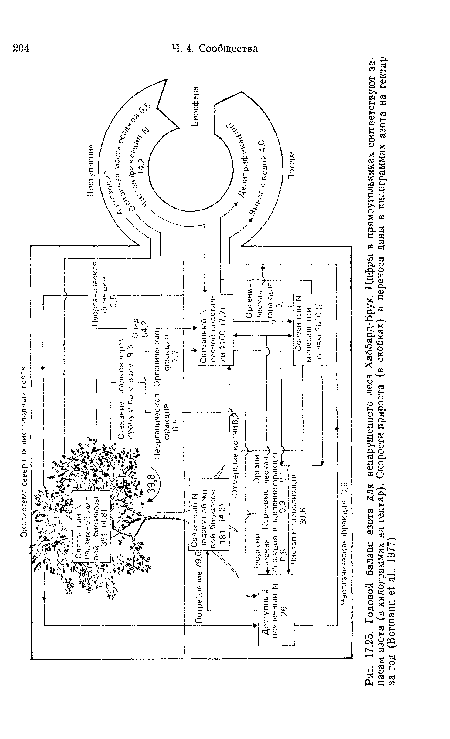 |
Далее
| Концентрация ионов для реки, в водосборном бассейне которой лес намеренно сведен, и для контрольного водотока (Хаббард-Брук). Момент сведения леса указан стрелкой. Отметьте наличие разрыва оси Y в случае нитрата (Likens, Bormann, 1975) |
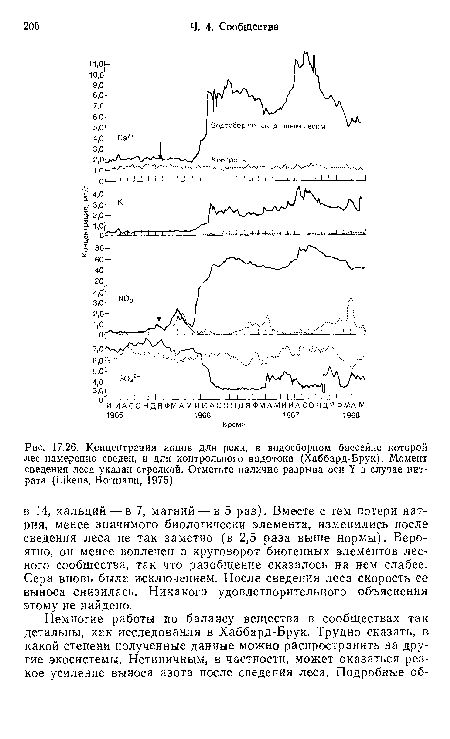 |
Далее
| Распределение азота по трем блокам органического вещества (надземному, корневому, почвенному) для шести типов биомов (Swift et al., 1979) |
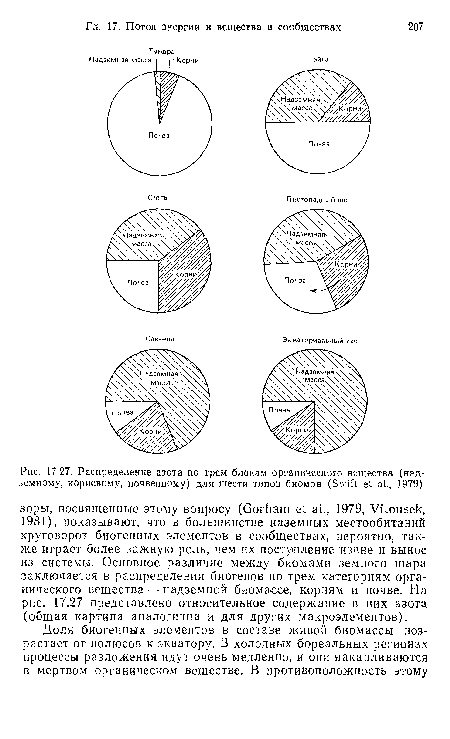 |
Далее
| Основные пути превращения азота в пресноводном озере (Moss, 1980) |
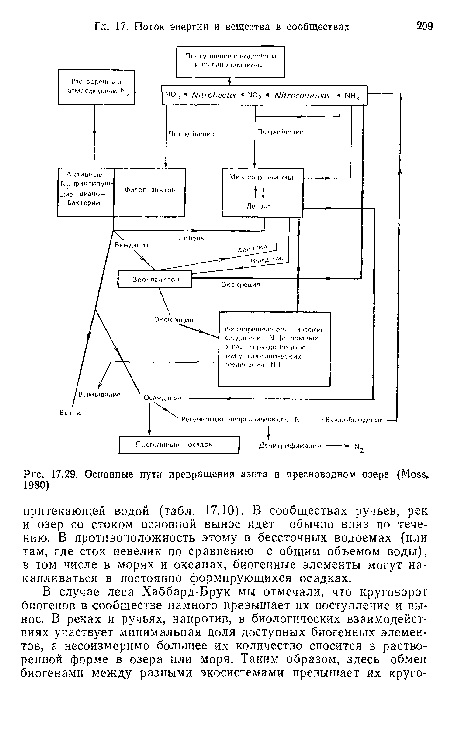 |
Далее
| Сосуществование конкурентов без расхождения ниш (компьютерное моделирование). Ниша вида 2 полностью входит в нишу вида 1, и в ходе конкуренции вид 2 теряет реализованную нишу. В однородной среде вид 1 (белые квадраты) полностью вытесняет вид 2 (белые кружки), однако, если среда мозаичная, а виды распределены отдельными скоплениями, они будут сосуществовать. Плотности популяций видов 1 (черные квадраты) и 2 (черные кружки) при этом ниже, чем в отсутствие межвидовой конкуренции (200 яиц у каждого). Сосуществование обусловлено не разделением ниш, а агрегированным распределением (по Atkinson, Shorrocks, 1981) |
 |
Далее
| Изменение доли разных видов шмелей вдоль вертикального траисек-та для двух длиннохоботных (Л) и трех короткохоботиых (Б) форм (Руке, 1982) |
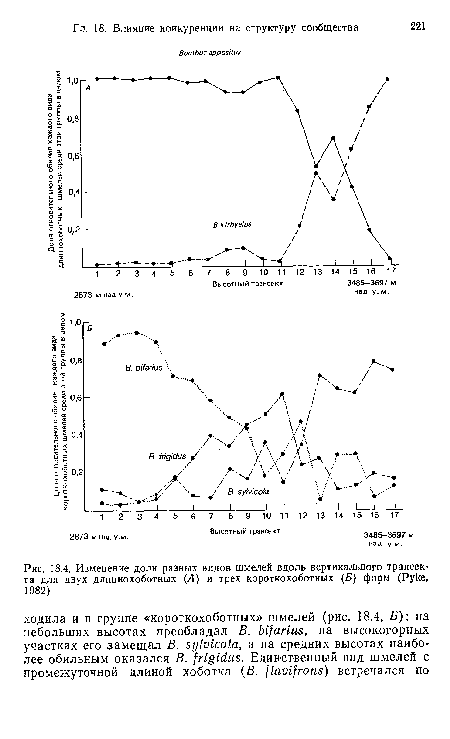 |
Далее
| Процент особей каждого вида бокоплавов рода Gammarus от общего их числа в пробах, взятых в пунктах с различной соленостью воды |
 |
Далее
| А. Последствия конкуренции между двумя видами за два ресурса будут зависеть от изоклин нулевого прироста каждого вида и положения точки снабжения этими ресурсами (см. разд. 7.11.1), Б. Результат конкуренции между пятью видами за два ресурса также зависит от положения изоклин нулевого прироста и точек снабжения ресурсами. Каждая отдельно взятая такая точка позволяет существовать либо одному виду, либо двум вместе, либо ни одному; однако в неоднородной естественной среде, включающей различные микросреды с широким набором этих точек (заштрихованная область), могут совместно обитать все пять (илн более) видов (по ТПтап, 1982, с изменениями) |
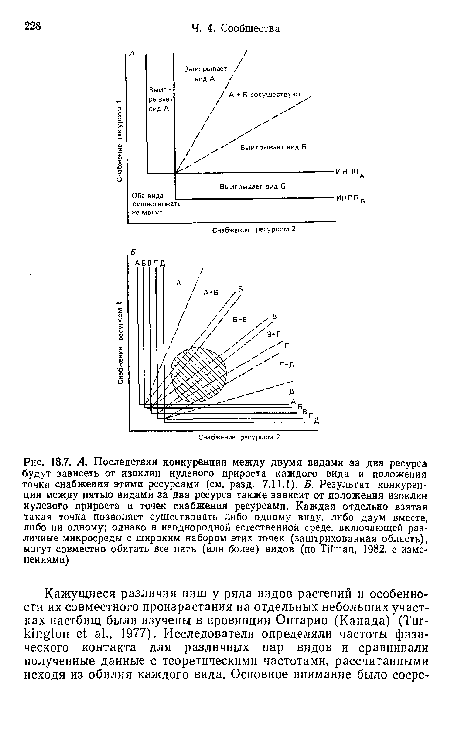 |
Далее
| Архипелаг Бисмарка, Соломоновы острова и Новые Гебриды, лежащие к востоку от Новой Гвинеи и Австралии |
 |
Далее
| Функции встречаемости для различных видов птиц архипелага Бисмарка. По оси ординат — доля островов, занятых данным видом, по оси абсцисс — «размер» острова (общее число живущих на нем видов птиц). |
 |
Далее
| Традиционный ансамбль музыкальных инструментов, размеры которых подчиняются правилу Хатчинсона (Horn, May, 1977) (инструменты любезно предоставил Russell Acott, Оксфорд; фото В. Roberts) |
 |
Далее
| Распределение размеров тела грызунов (использована логарифмическая шкала, так что равное расстояние между точками соответствует одинаковому отношению) в произвольно выбранных сообществах трех североамериканских пустынь. Заметно большое сходство распределения, хотя виды не всегда совпадают (Bowers, Brown, 1982) |
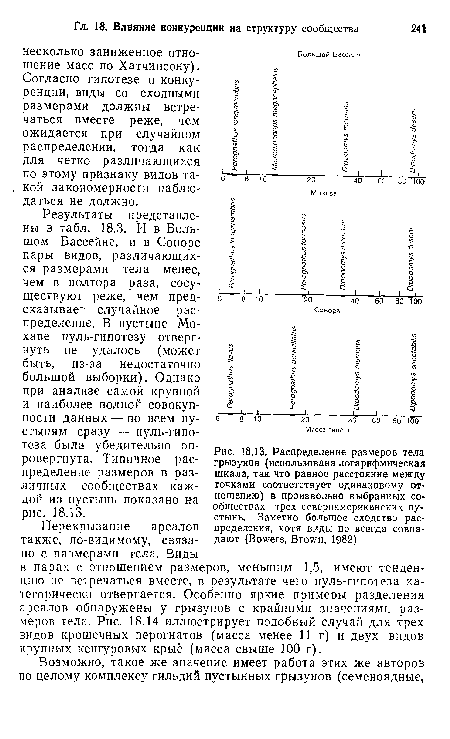 |
Далее
| Сравнение наблюдаемых (гистограммы) и полученных с помощью нейтральной модели (кривые) индексов сходства для попарного распределения видов птиц на архипелаге Бисмарка (Gilpin, Diamond, 1982). А. Все 11 325 пар видов при логарифмической шкале по оси ординат. Б. Шлейфы распределения при арифметической шкале по той же оси |
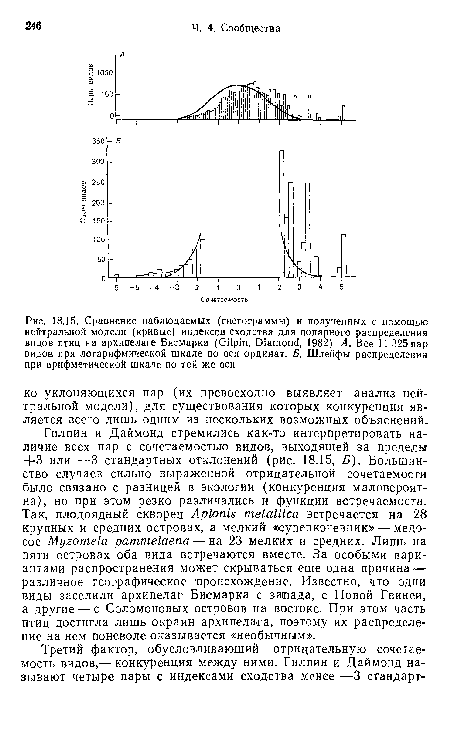 |
Далее
| Влияние плотности популяции улитки ЬШоппа ИЫогеа на видовое богатство (5) и разнообразие (индекс Шеннона, Я, рассчитанный на основе процента покрытия; с. 120) |
 |
Далее
| Видовое богатство фитопланктонного сообщества в целом (Л), съедобного (Б) и несъедобного (В) фитопланктона при низкой (белые кружки) и высокой (черные кружки) интенсивности выедания (по McCauley, Briand, 1979) |
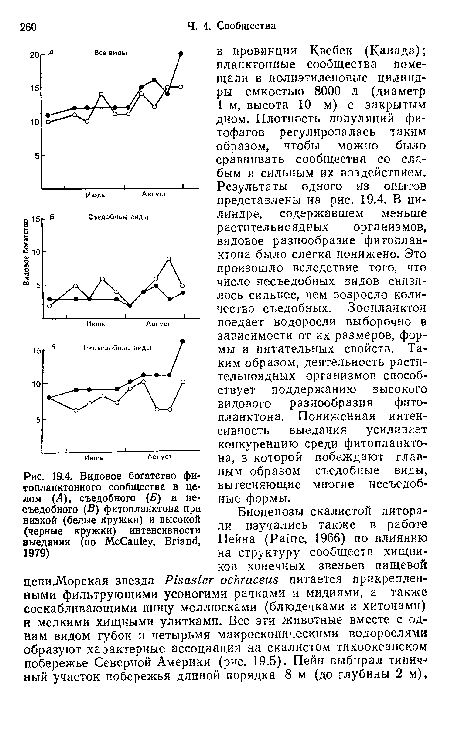 |
Далее
| Схема «конкурентной лотереи». Происходит захват периодически освобождающихся участков. Шансы на захват у всех видов (А—D) одинаковы н не зависят от видовой принадлежности предыдущего хозяина участка (это показано на врезке). Видовое богатство остается высоким и относительно постоянным |
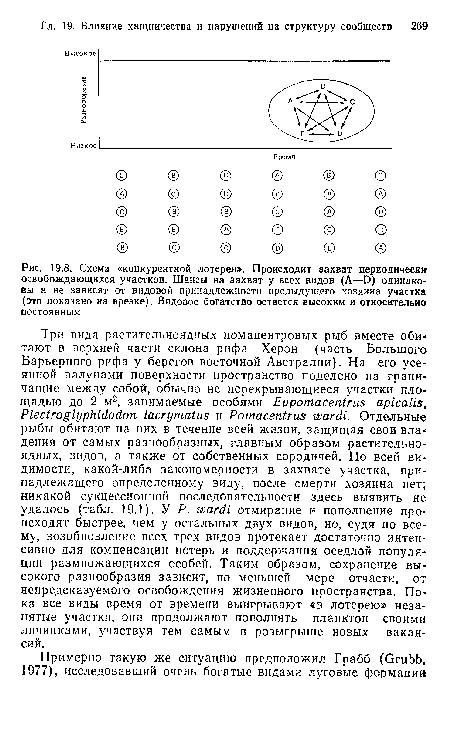 |
Далее
| Зависимость между температурой и вероятностью прорастания ряда однолетних и многолетних травянистых растений, обычных для лугов на известняках севера Англии (Grime, 1979) |
 |
Далее
| Кривые динамики видового богатства для трех освобождающихся время от времени участков н сообщества в целом при различных частотах нарушения. Прерывистые линии соответствуют стадии конкурентного исключения перед наступлением климакса |
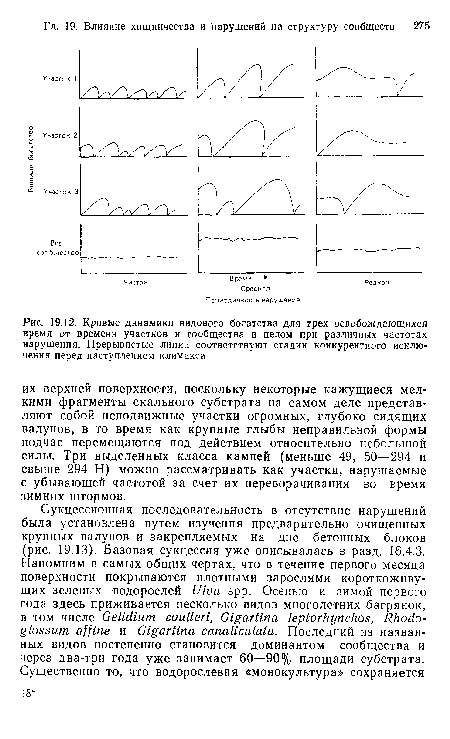 |
Далее
| Частотное распределение размеров участков в момент их образования на мидиевых банках в четырех пунктах северо-западного побережья Северной Америки (Paine, Levin, 1981) |
 |
Далее
| Моделирование исхода конкуренции между двумя видами. А. Среда постоянна н более благоприятна для вида Э]. Б. Среда остается более благоприятной для вида (случай I) или для вида Бг (случай II) на протяжении многих поколений; происходит конкурентное исключение одного вида, зависящее от момента начала взаимодействия. В. Среда более благоприятна для каждого вида по очереди в течение нескольких поколений (время, необходимое для конкурентного исключения, примерно равно требующемуся для изменения среды); в этих условиях возможно сосуществование |
![Моделирование исхода конкуренции между двумя видами. А. Среда постоянна н более благоприятна для вида Э]. Б. Среда остается более благоприятной для вида (случай I) или для вида Бг (случай II) на протяжении многих поколений; происходит конкурентное исключение одного вида, зависящее от момента начала взаимодействия. В. Среда более благоприятна для каждого вида по очереди в течение нескольких поколений (время, необходимое для конкурентного исключения, примерно равно требующемуся для изменения среды); в этих условиях возможно сосуществование](/static/pngsmall/228045472.png) |
Далее
| Изменения в видовом составе заливного луга в пойме Рейна (ФРГ) в зависимости от интенсивности затопления и отложения ила в разные годы (по Müller, Foerster, 1974) |
 |
Далее
| Влияние неравновесных условий на исход конкуренции. А. Моделирование ситуации с конкурентным исключением. Б. Конкурентное исключение предотвращается периодическим, не зависящим от плотности сокращением популяций (по Huston, 1979) |
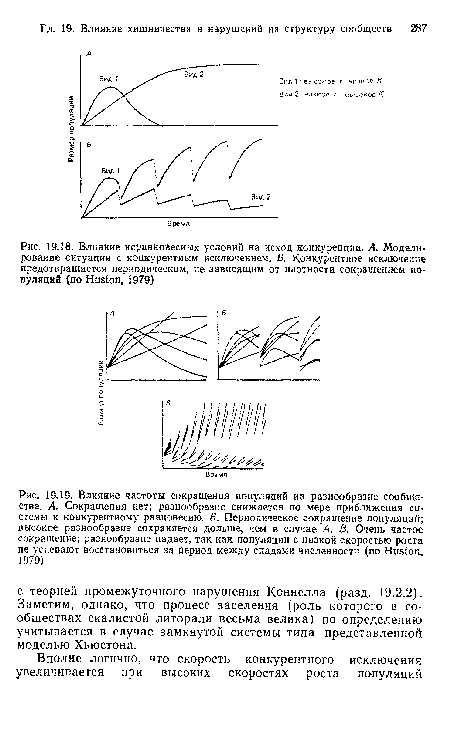 |
Далее
| Результаты моделирования сообществ по Хьюстону. А. Ожидаемая зависимость между разнообразием и частотой сокращения популяций. Б. Ожидаемая зависимость между разнообразием и скоростью роста популяций в неравновесных системах при низких и средних частотах сокращений численности. В. Обобщенный график зависимости между скоростью конкурентного вытеснения (эквивалент скорости роста популяций), частотой сокращения популяций и видовым разнообразием. Разнообразие изменяется по оси, перпендикулярной плоскости страницы, и представлено в виде замкнутых кривых (по Huston, 1979) |
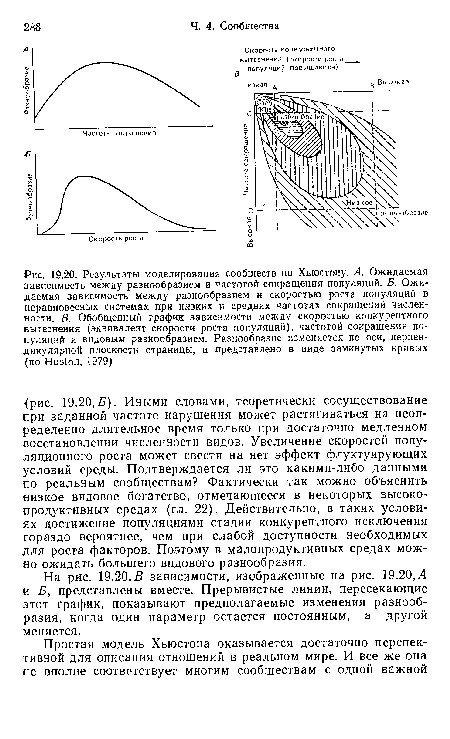 |
Далее
| График зависимости между числом видов цветковых растений и площадью для учетных площадей различных размеров в Англии (Williams, 1964, из Gorman, 1979) |
 |
Далее
| Равновесная теория островной биогеографии Мак-Артура и Уилсона (MacArthur, Wilson, 1967). А. Зависимость скорости иммиграции от числа обосновавшихся видов для крупных и мелких островов и для островов, лежащих вблизи и вдали от континента. Б. Зависимость скорости вымирания от числа обосновавшихся на острове видов для крупных и мелких островов. В. Баланс между иммиграцией и вымиранием для крупных и мелких и для близких и удаленных от континента островов. В каждом случае 5 обозначает равновесное видовое богатство. Подробности см. в тексте |
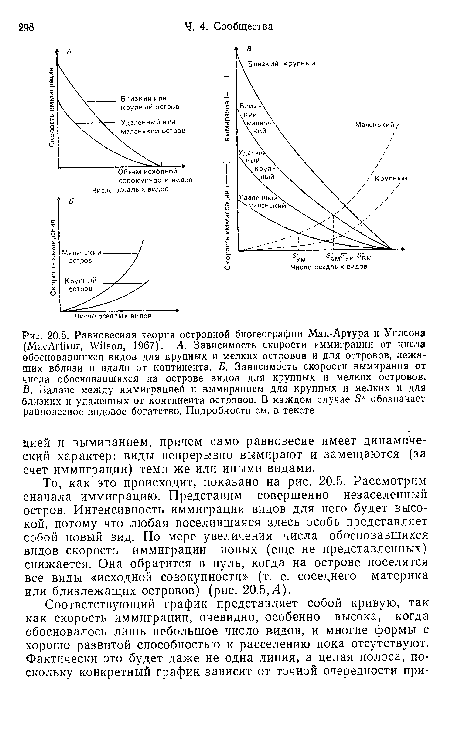 |
Далее
| Влияние искусственного сокращения площади мангровых островов на число видов членистоногих. Размеры островов 1 и 2 были сокращены после учетов 1969 и 1970 гг., острова 3 — только после учета 1969 г. Изменение видового богатства на контрольном острове вызвано, по-видимому, случайными флуктуациями (по БтЬегЫ^ 1976) |
 |
Далее
| Зависимость видового богатства ящериц иа островах Калифорнийского залива, некогда бывших частями сухопутного перешейка, от продолжительности их изоляции (по Wilcox, 1978) |
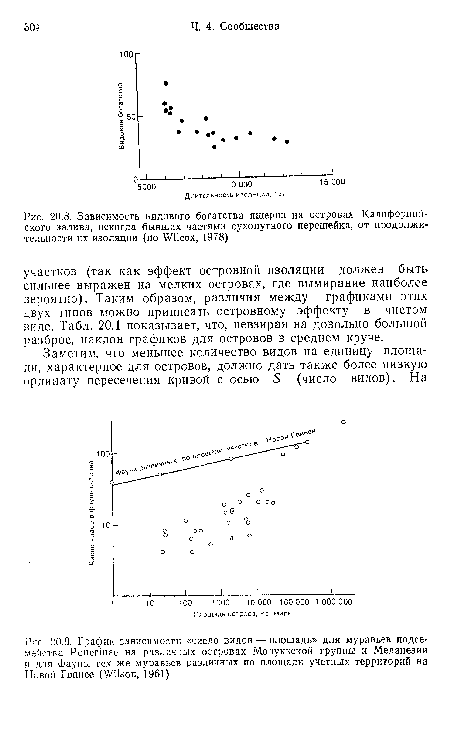 |
Далее
| График зависимости «число видов — площадь» для муравьев подсемейства Ponerinae на различных островах Молуккской группы и Меланезии и для фауны тех же муравьев различных по площади учетных территорий на Новой Гвинее (Wilson, 1961) |
|
Далее
| Связь числа неморских равнинных видов птиц островов, расположенных на расстоянии свыше 500 км от Новой Гвинеи (в процентах от числа видов на острове сходной площади у ее берегов), с удаленностью этого источника орнитофауны (Diamond, 1972) |
 |
Далее
| Динамика роста видового богатства различных групп бентосной фауны залива Тампа после естественной гибели животных (по Simon, Dauer, |
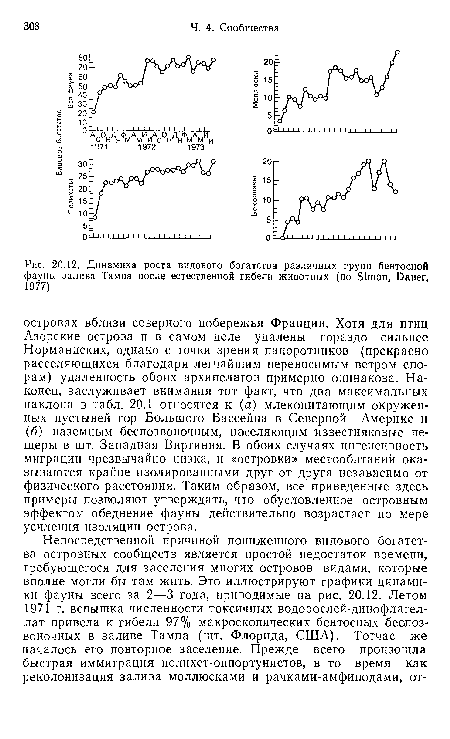 |
Далее
| Отношение между числом видов и площадью для растительноядных насекомых на орляке. В тех регионах, где этот папоротник распространен шире, на нем обитает больше видов. Линия регрессии построена по черным кружкам (белые кружки соответствуют учетам, включающим нерегулярно или случайно попадающиеся виды) (Lawton, 1984) |
 |
Далее
| Видовое богатство насекомых-фитофагов на растении возрастает по мере усложнения его архитектоники. А. Отношения «число видов-—площадь» для разных рода деревьев, кустарников и трав британской флоры; ареал кормового растения представлен как количество квадратов со стороной 10 км на карте (Strong, Levin, 1979). Б. Зависимость числа видов, связанных с разными видами рода Opuntia в Северной и Южной Америке, от «архитектурной сложности» этих кактусов, выражаемой суммой значений пяти параметров, каждый из которых оценивается по четырехбалльной шкале |
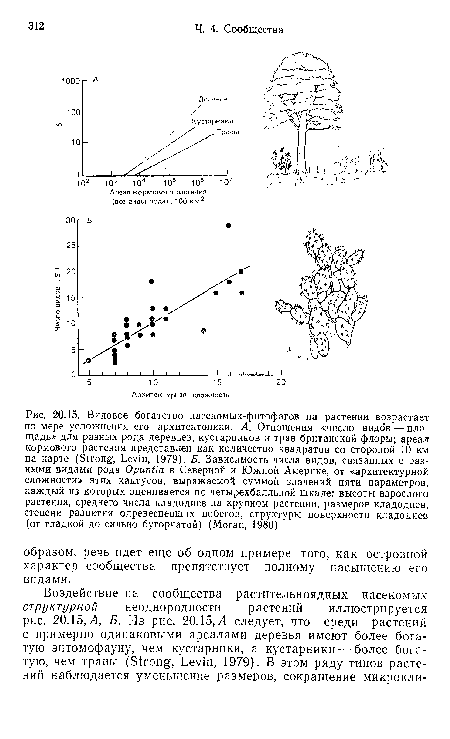 |
Далее
| Восточные границы распространения семейств и подсемейств гнездящихся наземных и водоплавающих пресноводных птиц, встречающихся на Новой Гвинее. Сокращение числа таксонов происходит постепенно и отражает как различные способности групп к расселению, так и общее уменьшение размеров островов с запада на восток. 1 — не встречаются за пределами Новой Гвинеи |
 |
Далее
| В небольшом дубовом лесу Истерн-Вуд на юге Англии одни виды» птиц никогда не исчезают, а другие периодически эмигрируют и вселяются заново. Частотная гистограмма показывает число эмиграций с последующим возвращением за 26 лет. На каждый вид приходится в среднем 3,4 случая (по-Веуеп, 1976) |
 |
Далее
| Относительное видовое богатство членистоногих-фитофагов, связанных с тремя древесными породами в Великобритании (заштриховано) и Южной Африке. В каждом случае с интродуцированным растением (И) связано меньше видов, чем с местным (М) (по БоиНшоо!! е1 а!., 1982) |
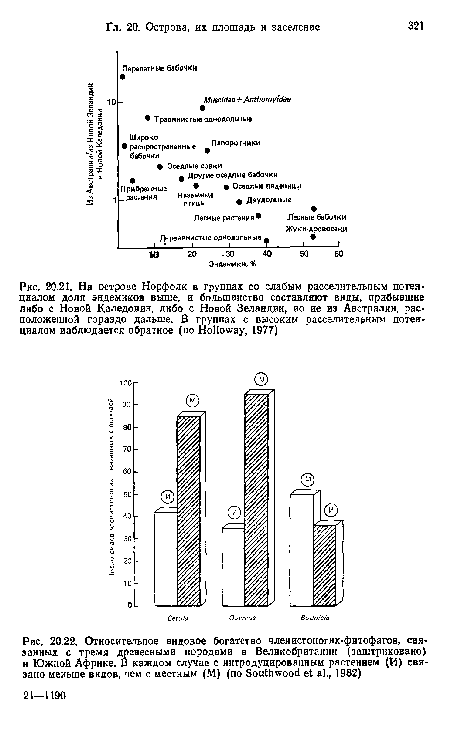 |
Далее
| Наглядные аналогии, иллюстрирующие различные аспекты устойчивости, используемые в этой главе для описания сообществ |
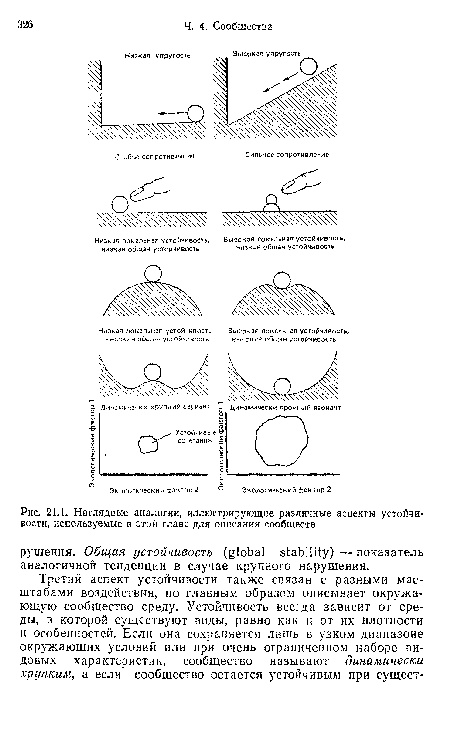 |
Далее
| Сложность и упругость модельных пищевых сетей. По мере роста сложности (в данном случае —связности) возрастает доля устойчивых пищевых цепей с малым временем релаксации (Р) после нарушения (быстрая релаксация свидетельствует об упругости). Кружки — модели с тремя трофическими уровнями, треугольники — с четырьмя (по Pimm, 1982) |
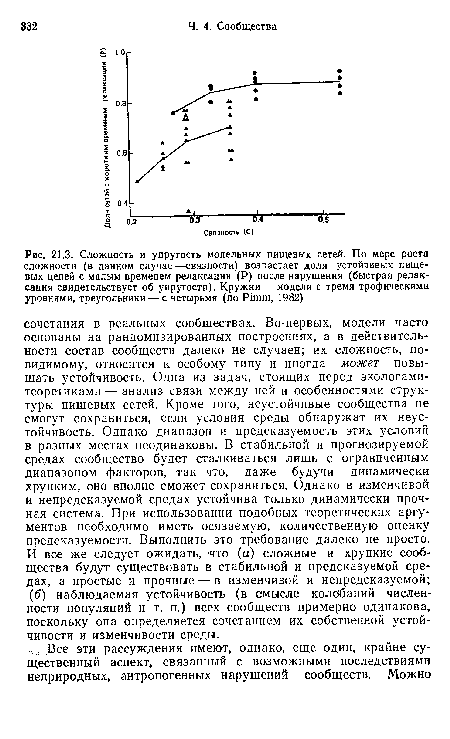 |
Далее
| Важнейшие взаимодействия внутри трех взаимосвязанных местообитаний о. Медвежий (Северный Ледовитый океан) и между ними |
 |
Далее
| Расчлененность пищевой сети. Перемещения метки 32Р из растений двух видов в залежном сообществе. Цифры в кружках |
|
Далее
| Гистограммы частотного распределения средних длин максимальных пищевых цепей в сообществах (Л) изменчивой и (£>) постоянной сред. Общие средние значимо ие различаются (3,66 звеньев в первом случае и 3,60 — во втором) (по данным Впаш1, 1983) |
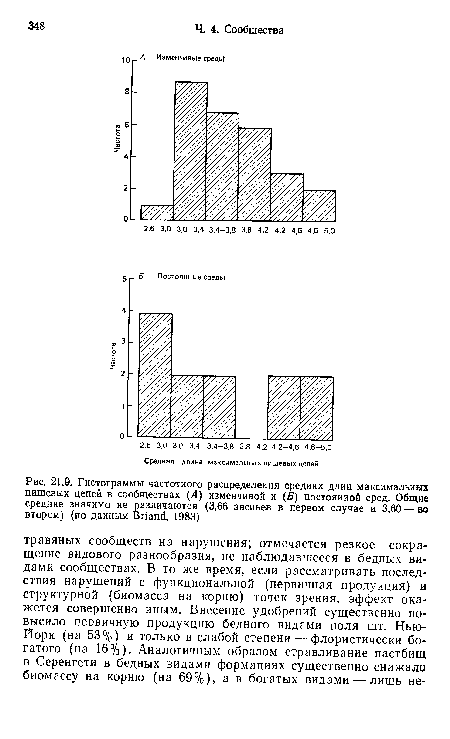 |
Далее
| Простая модель сообщества. Три блока соответствуют компонентам системы, а стрелками обозначен перенос энергии между ними (O’Neill, 1976) |
 |
Далее
| Зависимость скорости восстановления (показатель упругости) после нарушения от притока энергии на единицу биомассы иа корню для моделей шести различных сообществ. Наиболее упругой системой оказался пруд, наименее—тундра (по данным O’Neill, 1976) |
 |
Далее
| Простая модель видового богатства. Каждый вид использует долю п доступного ресурса Я; перекрывание с соседними видами составляет о. В одном сообществе может быть больше видов, чем в другом, из-за того, что |
 |
Далее
| Более продуктивная среда способна обеспечить существование большего числа более специализированных видов (п при равновесном состоянии меньше) |
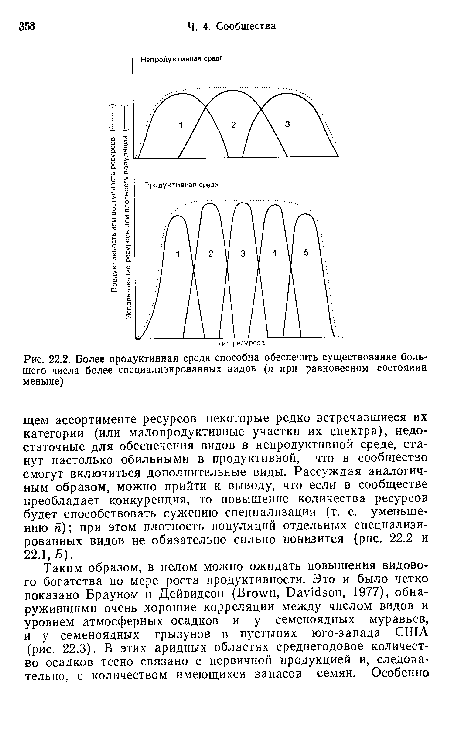 |
Далее
| Связь видового богатства семеноядных грызунов (треугольники) и муравьев (кружки) на песчаных почвах с географическим градиентом выпадающих осадков и продуктивности (по Brown, Davidson, 1977) |
 |
Далее
| При возрастании продуктивности видовое богатство может сокращаться. Снабжение двумя ресурсами (1 и 2) позволяет сосуществовать пяти видам (А—Д), согласно работе Тилмана (ТПтап, 1982), рассмотренной в гл. 18 (рис. 18.7,Б). Темный круг внизу слева показывает снабжение ресурсами в относительно малопродуктивной среде, позволяющее сосуществовать всем пяти видам. Темный круг вверху справа соответствует относительно высокопродуктивной среде (но с таким же диапазоном различных значений снабжения); здесь могут сосуществовать лишь два вида |
 |
Далее
| Зависимость между |
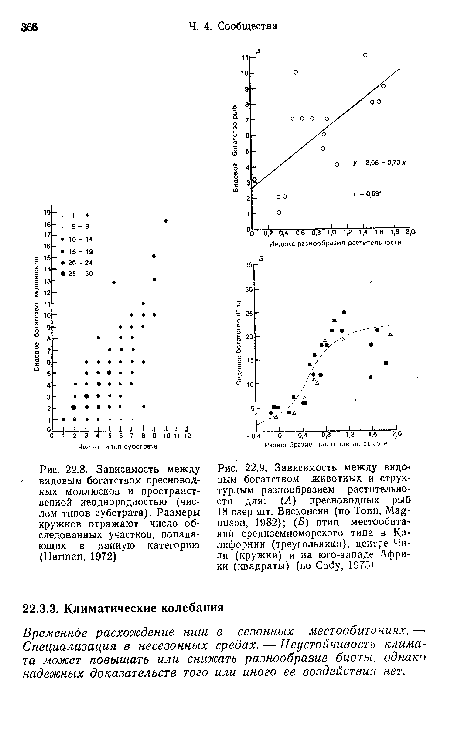 |
Далее
| Зависимость между видовым богатством и диапазоном перепадов температуры в различных точках западного побережья Северной Америки для (А) птиц, (Б) млекопитающих, (В) брюхоногих моллюсков (по МасАгШиг, 1975) |
 |
Далее
| Широтное распределение видового богатства |
 |
Далее
| Приживаемость всходов дерева дождевых тропических лесов Dipte-ryx panamensis возрастом от 7 месяцев до 2 лет в зависимости от расстояния до ближайшей взрослой особи (Clark D. A., Clark D. В., 1984) |
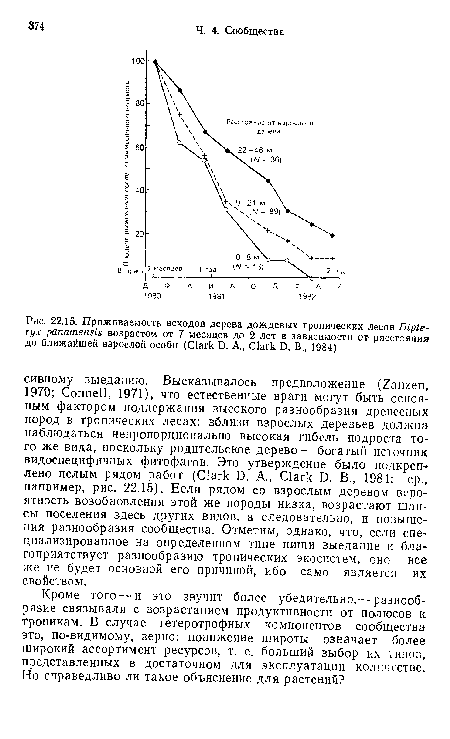 |
Далее
| Суммарное число видов жуков семейств Scolytidae и Platypodidae по категориям пищевой специализации. В высоких широтах специализация этих насекомых, как правило, уже (данные из Beaver, 1979) |
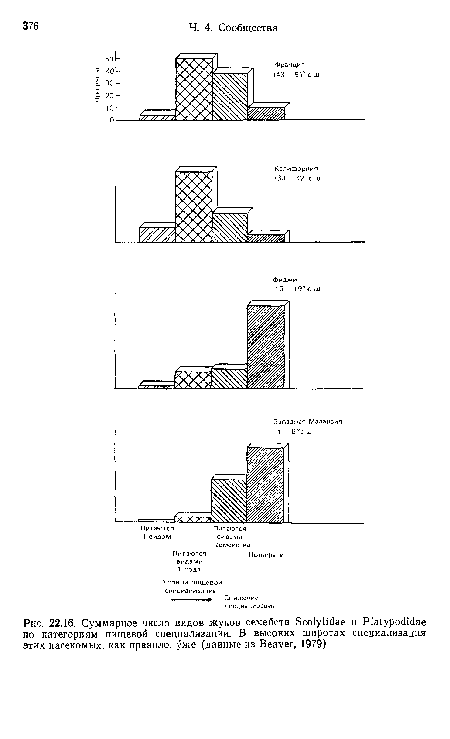 |
Далее
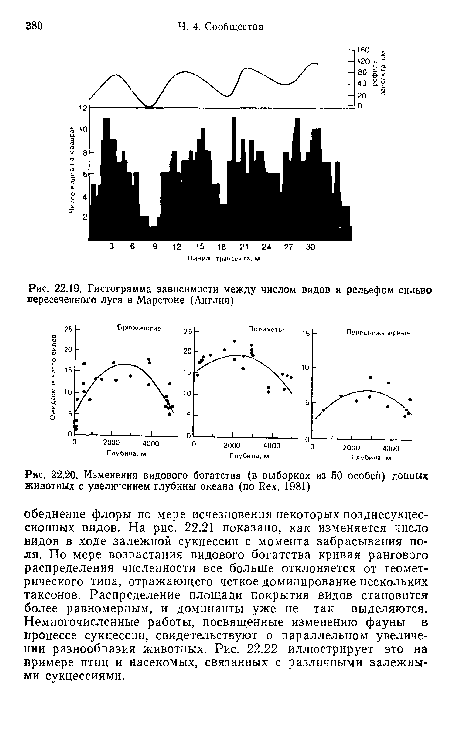
| Гистограмма зависимости между числом видов и рельефом сильно пересеченного луга в Марстоне (Англия) |
 |
Далее
| Изменения видового богатства (в выборках из 50 особей) донных животных с увеличением глубины океана (по Rex, 1981) |
|
Далее
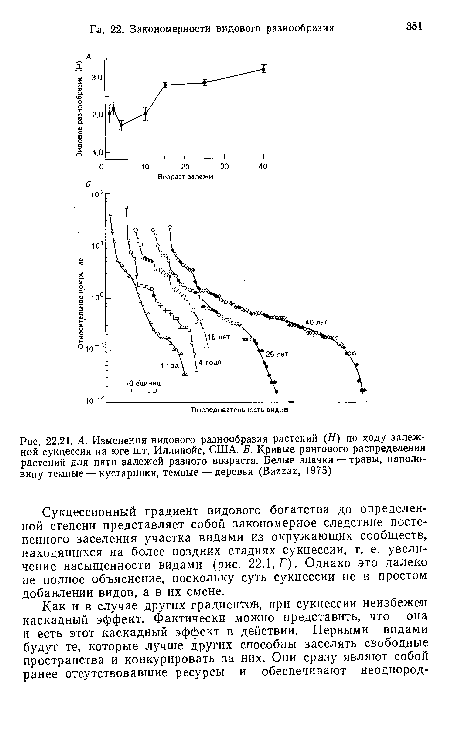
| А. Изменения видового разнообразия растений (Я) по ходу залежной сукцессии иа юге шт. Иллинойс, США. Б. Кривые рангового распределения растений для пяти залежей разного возраста. Белые значки — травы, наполовину темные — кустарники, темные — деревья (Ваггаг, 1975) |
 |
Далее
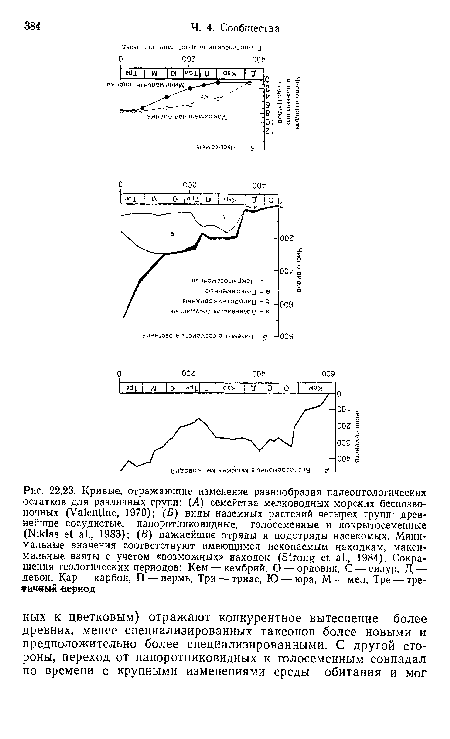
| Кривые, отражающие изменение разнообразия палеонтологических остатков для различных групп |
 |
Далее
| Число видов наземных животных по классам размеров тела (длины). Знак вопроса означает, что это чисто прикидочная гистограмма, основанная на самых грубых оценках (May, 1978) |
 |
Далее