Гаметогенез - процесс формирования половых клеток (гамет).[ ...]
Начало гаметогенеза - это самая продолжительная стадия полового цикла. Она отмечалась с середины октября до середины мая. На ее протяжении можно отметить довольно резкое увеличение за счет роста ацинусов. Половые железы окрашены в слабо-фиолетовый цвет. Гонады приобретают некоторый тургор по сравнению с посленерестовой стадией. В начале стадии ацинусы очень малы. Ооциты располагаются вдоль ацинусов, соединяясь с ними широким основанием.[ ...]
Акимова Н.В. Гаметогенез и половая цикличность сибирского осетра в естественных и экспериментальных условиях // Особенности репродуктивных циклов у рыб в водоемах разных широт. М.: Наука, 1985.[ ...]
Особенности гаметогенеза окуневых (сем, РегсМае) и карповых (сем.[ ...]
А. Н. Гаметогенез и сравнительный анализ развития воспроизводительной системы у пеляди, выращиваемой в разных климатических зонах.— Изв..[ ...]
Стадия активного гаметогенеза отмечалась с конца мая по июнь. Половые железы становятся плотными, окрашенными в темно-фиолетовый цвет. В ацинусах яичников можно встретить генерации половых клеток от гониев до ооцитов, закончивших рост, диаметром 60-100 мкм.[ ...]
Г. Изучение связей гаметогенеза с особенностями обмена веществ у особей различных видов рыб (эколого-физиологическое направление исследований).[ ...]
Макроспорогенез и гаметогенез у них составляют единую цепь клеточных делений, завершающим звеном которой является формирование женского гаметофита крайне упрощенного строения, превратившегося во внутренний орган спорофита. Развитие его максимально сокращено и структура доведена до нескольких клеток. Однако несмотря на морфологическую редукцию, зародышевый мешок состоит из обособленной системы клеток, отличающихся четкой функциональной дифференциацией на разных этапах их развития.[ ...]
М.: ИЭМЭЖ АН СССР, 1978.[ ...]
Пути и формы адаптации гаметогенеза и половых циклов у пресноводных окуневых рыб к различным условиям существования. Подытоживая изложенные данные по гаметогенезу пресноводных видов рыб семейства Percidae, следует отметить, что представители одного семейства, даже близкородственные виды, как ерш и носарь, относящиеся к одному роду ершей — Acerina, имеют различную биологию размножения — это и экология нереста, и специфика развития и выметывания половых клеток.[ ...]
Таким образом, изучение гаметогенеза, половой цикличности, ритма размножения, экологии нереста и особенностей воспроизводства у некоторых видов сиговых рыб было необходимым звеном в наших сравнительных эколого-морфологических исследованиях различных сторон репродуктивного процесса у рыб.[ ...]
Кошелев Б. В. Роль темпов гаметогенеза и овариальных циклов в воспроизводстве популяций у видов рыб в различных условиях существования.— В кн.: Эволюция темпов индивидуального развития животных. М,: Наука, 1977а, с. 186—199.[ ...]
На VII этапе идут процессы гаметогенеза, формируется пыльца. На VIII этапе заканчивается формирование зародышевого мешка и яйцеклетки.[ ...]
У самок сига при достижении сроков первого нереста, как и у других видов рыб, период превителлогенеза, период ядерно-плаз-менных преобразований, развивающихся ооцитов, может протекать в течение весьма разного отрезка времени, что определяет ту или иную скороспелость особей.[ ...]
Роль регулятора процесса гаметогенеза в условиях изменения режима освещения отводят пинеальному органу, или эпифизу, который, видимо, является у рыб не просто светочувствительным рецептором нервной системы (Omura, Oguri, .1970), но, подобно эпифизу у более высокоорганизованных животных, выполняет роль эндокринного секреторного органа (Friederick-Freska, 1932; Pflugfelder, 1953, 1954; Rüderberg, 1968; Fenwick, 1970).[ ...]
Персов Г. М. Ранний период гаметогенеза у проходных лососей.— Тр.[ ...]
Питание в регуляции цикла гаметогенеза у разных видов рыб играет неодинаковую роль. У угря Anguilla anguilla, например, протекание сперматогенеза не зависит от условий питания. Самцы его нормально созревают даже после трехлетнего периода голодания (Boétius, Boétius, 1967). Золотой карась, напротив, быстро реагирует на недостаток пищи (Clemens, Reed, 1967). В течение четырех месяцев ограниченного кормления в период, соответствующий естественному нересту, относительный вес семенников падает у него в десять раз. Сперматогенез удается восстановить введением препаратов гипофиза в сочетании с повышением суточного рациона.[ ...]
Исследования особенностей гаметогенеза и прохождения полового цикла у сибирского осетра, которые проводятся под нашим руководством Акимовой, сочетаются с анализом динамики содержания жира в разных органах в течение всех стадий зрелости половых желез, на протяжении всей жизни особей. Выяснение же-изменений в характере гаметогенеза, скорости полового созревания и ритма размножения, наряду с темпом роста и уровнем метаболизма у акклиматизированного сибирского осетра в ряде водоемов европейской части СССР, дает возможность судить уже не о потенциальных, а о реальных адаптивных возможностях данного вида, о путях и способах приспособления различных звеньев репродуктивного процесса, а также отдельных этапов и периодов онтогенеза к различным условиям существования (см. рис. 80).[ ...]
Дормидонтов А. С. Особенности гаметогенеза сигов в северных водоемах Якутии.— Там же, 1974а.[ ...]
Глубокое влияние оказывают на гаметогенез различные виды излучений. Семенники гуппи довольно быстро реагируют на облучение.[ ...]
Для выяснения закономерностей гаметогенеза, форм и путей приспособления всех звеньев репродуктивного процесса были выбраны не только различные экологические формы сига, но и близкородственные виды среди окуневых, карповых и других семейств, обитающие в пределах, участках.ареала, а также различные формы окуня (прибрежный и пелагический), карася (обыкновенный золотой карась и низкотелая тугорослая его форма), проходной и турв водный лещ, разные по темпу роста формы воблы, красноперка и др. виды рыб [Кошелев, 1961а, б, в, 1962, 1965а, б, 1968, 1970; Шихшабеков, 1969].[ ...]
| Распределение хромосом при гаметогенезе | 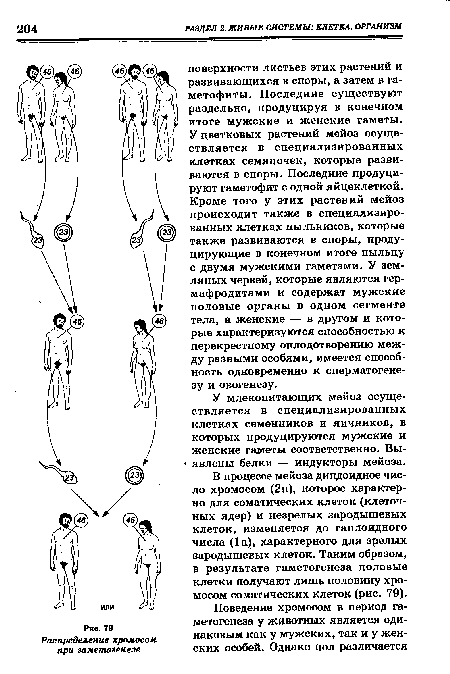 |
Расщепление генов происходит при гаметогенезе во время мей-отических делений. При мейозе хромосомы расходятся и проходят в разные гаметы, причем каждая гамета получает по одной хромосоме (гомологу) из пары хромосом. Независимое распределение генов также может быть объяснено поведением хромосом при мейозе (рис. 127).[ ...]
Главным признаком нарушения гаметогенеза являлась асин-хронность созревания яйцеклеток.[ ...]
Кошелев Б, В, Изучение размножения рыб (гаметогенез, скорость полового созревания, половая цикличность, ритм икрометания и экология нереста).—■ В кн.; Исследования размножения и развития рыб: (Метод, пособие). М.: Наука, 19816, с. 5—16.[ ...]
Л,: ЛГУ, 1964.[ ...]
Кошелев Б. В. Исследование закономерностей гаметогенеза и половых циклов у рыб в различных частях ареала.— В кн.: Типовые методики исследования продуктивности видов рыб в пределах их ареалов, ч. I. Вильнюс: Минтис, 1974, с. 37—44.[ ...]
Д. Анализ закономерностей прохождения и изменения гаметогенеза, половых циклов и ритма размножения у разных видов рыб в связи с разнообразными условиями существования (эколого-мор-филогическое направление исследований).[ ...]
Кошелев Б. В. Эколого-морфологическое исследование гаметогенеза, половой цикличности и размножения рыб.— В кн.: Эколого-морфологические и эколого-физиологические исследования развития рыб. М.: Наука, 1978, с. 10—42.[ ...]
Головка спермия представляет собой (преобразованное в процессе гаметогенеза ядро ш ер м а тог он и а л ь но й клетки и состоит в основном из дезоксирибонуклеопротеида (ДНП). У черноморско-азовского осетра Acipenser güldenstàdti colchi-cus в спермиях на долю ДНП приходится 90,5%, а у лосося Salmo sedar — 98,1% вещества головки (Георгиев и др., 1960; Збарский, Ермолаева, 1961; Ермолаева, 1964). Здесь содержится материал гаплоидного набора -хромосом, в силу чего абсолютное количество ДНК в головке спермия вдвое ниже, чем в ядрах соматических клеток (Mirsky, Ris, 1949, и др.).[ ...]
У ряда видов покрытосеменных растений микроспоры в течение всего гаметогенеза остаются соединенными в тетрады (Вересковые, Ситниковые и др.) или более крупные группы, называемые поллиниями (Орхидные, Мимозовые), т. е. образуют сложную пыльцу.[ ...]
Для некоторых видов рыб эффективными раздражителями, влияющими на гаметогенез, являются: течение (Державин, 1947; Кузьмин, Чуватова, 1970) и изменение уровня воды (Ноаг, 1959; John, 1963; Hyder, 1969), условия питания, насыщение воды кислородом (Строганов, 1952; Clemens, Reed, 11967), присутствие особей противоположного пола (Фалеева, 1958; Egami, 1959; Amouriq, 1964) и др. Особенно заметно на развитие воспроизводительной системы влияют температура и свет (Hyder, 1969; Nomura, 1969; Schneider, Immelmann, 1969; Schneider, 1969; Wiebe, 1969b). Степень воздействия каждого из перечисленных факторов зависит от особенностей биологий вида, а также физиологического состояния особи в момент воздействия, стадии ее развития или этапа гаметогенеза и т. д. Важную роль играет определенное сочетание, комплексное действие многих раздражителей, приводящее к реализации механизма управления половым циклом (Поликарпова, 1942; Строганов, 1962, и др.).[ ...]
Пополнение запасов половых клеток у половозрелых самцов на протяжении соответствующего цикла гаметогенеза в большинстве случаев описывается как интенсивный процесс митотических делений покоящихся клеток, которые постоянно присутствуют в стенках семенных трубочек, являются прямыми потомками первичных половых клеток и в определенный период активируются к дальнейшему развитию. Их называют покоящимися — dormant germ-cells (Hann, 1927), резервными— reserve germ-cells (Suzuki, 1939), остаточными — resting germ-cells (Jones, 1940; Weisel, 1943), остаточными зачатковыми семенными — resting sperm mother cell s (Lehri, 1967; Rai, 1967) или зачатковыми (Кулаев, 1928, и др.). Каждое из названий характеризует одно из свойств этих клеток, которые являются одновременно и зачатковыми, и покоящимися, и остаточными, и резервными.[ ...]
Вопросы созревания и формирования гамет у корбикулы японской, обитающей в зал. Петра Великого, не изучены. В связи с этим в данной работе исследованы гаметогенез и репродуктивный цикл этого моллюска.[ ...]
Особенности развития половых клеток у eptua, связанные с асинхронным ростом ооцитов и порционным типом икрометания. Для выяснения особенности сезонного хода гаметогенеза и закономерностей ритмичного развития яйцеклеток в половых железах особей, населяющих водоемы разных широт, был выбран близкий к окуню вид — ерш (Acerina cernua L.). По многочисленным литературным данным, ерш имеет порционный тип икрометания. Нерестовый период длится в течение нескольких месяцев и, как правило, наблюдается в меженный период [Лукин, 1948а, б; Васнецов, 1949; Дрягин, 1949а, б; Толчанов, 1951; Шмидтов, Варфоломеев, 1952; Крыжановский и др., 1953; Буцкая, 1955, 1957, Í959; и др.]- Несмотря на то что ерш за последнее время стал занимать одно из ведущих мест в ихтиофауне и в промысловых уловах во многих реконструированных и в некоторых природных водоемах, его биология размножения, и особенно гаметогенез и половая цикличность, еще мало изучены. Благодаря широкому распространению (см. рис. 22), обилию ерша во многих водоемах он вылавливался самыми различными орудиями лова: мутниками, неводами, сетями, подъемниками и разного рода любительскими орудиями лова (сачками, переметами и удочками). Благодаря применению разнообразных орудий лова мы имели ершей в любое время года в достаточном количестве.[ ...]
Итак, генные пары двух скрещиваемых организмов расщепляются (сегрегируют) независимо одна от другой и перераспределяются, подвергаясь свободной рекомбинации между собой, причем гибридные организмы в процессе их гаметогенеза дают начало комбинациям родительских гамет и рекомбинациям гамет в равных количествах. Данное правило, установленное Г. Менделем, в настоящее время называют вторым законом наследственности (вторым законом Г. Менделя), законом независимого перераспределения генных аллелей или законом рекомбинации генов.[ ...]
Несмотря на то что, по-видимому, судак стоит ближе, чем окунь и другие виды, к исходной форме, от которой произошли все пресноводные окуневые, у него на относительно обширном ареале обнаруживаются различного рода изменения в гаметогенезе и половых циклах, однако никогда не приводящие к порционному типу икрометания. Эти особенности в скорости и в характере развития половых клеток и сезонном ходе гаметогенеза являются адаптацией к различным условиям существования и тесно связаны с уровнем и спецификой обменных процессов у особей в различных водоемах.[ ...]
Половое созревание сига-пыжьяна растянуто на ряд лет, и для него характерны пропуски в икрометании после первичного полового созревания (Москаленко, 1955, 1958а, 1971; Дор-мидонтов, 1974, 1974а, и др.). Ранее некоторыми исследователями уже была показана зависимость гаметогенеза и нерестовых миграций у этого и близких к нему видов от экологических условий (Решетников, 1966, 1967; Решетников и др., 1971). В пределах обследованного нами бассейна эта рыба впервые созревает в возрасте 3+- лет, а основная часть ее особей — в 5+ и 6+ лет. В августе — сентябре на р. Ляпине, в сентябре — октябре на ее притоках поднимающийся на нерест и-последую-щую зимовку сиг-пыжьян был представлен исключительно половозрелыми особями с гонадами в IV стадии зрелости.[ ...]
Длительность митоза первичного ядра в пыльцевом зерне зависит от условий среды (температура, влажность воздуха), характера питания микроспор и особенностей развития тапе-; тума пыльника. Так, в пыльцевых зернах традесканции длительность отдельных фаз митоза в период гаметогенеза следующая (при температуре 30°С): профазы — 30—34 ч (наибольшая часть этого времени приходится на раннюю профазу), метафазы и анафазы — по 0,5, телофазы — 0,2 ч. Весь митоз длится от 31,2 до 35,2 ч (Бкшоп, 1950).[ ...]
Наши дальнейшие исследования показали значительную стерильность М 2 по мужской линии. У нее только часть пыльцы бывает фертильной, причем количество ее варьирует от 0 до 80%. Специальным экспериментом выявлено, что это варьирование в большой степени зависит от температуры и влажности среды во время макроспорогенеза, гаметогенеза и цветения. Умеренная температура и высокая влажность способствуют формировании» нормальной пыльцы и растрескиванию пыльников. У отдельных, растений, развивающихся при одинаковых условиях, несколько отклоняется соотношение стерильной и фертильной пыльцы а также процент растрескивающихся пыльников.[ ...]
В настоящее время сводки, которые охватывали бы круг вопросов, рассматриваемых в книге, отсутствуют. Существенную трудность при составлении такого обзора исследований представляет то, что интересующие нас материалы публикуются в чрезвычайно большом количестве источников самого различного профиля. В известной мере облегчили нашу задачу обзорные работы по вопросам происхождения и миграции первичных половых клеток (Vivien, 1964; Персов, 1969; А. Тур-даков, 1969а), дифференцировки пола (Юровицкий, 1966; Кирпичников, 1969; Персов, 1969), гермафродитизму (Салехова, 1965, 1970; Персов; 1969), гормональной регуляции гаметогенеза (Bern, 1967; Lofts, 1968; Qordt, 1968; Ванякина, 1969), строению и свойствам спермиев рыб (Гинзбург, 1968).[ ...]
Цветение растений АД2 первой группы очень растянуто, так как у каждого растения имеются побеги разного возраста. В основном цветение в 1972 г. совпало с жаркой и сухой погодой, а в 1973 г.— с переменной температурой и влажностью, у большинства растепий цветки во время цветения были открыты, как у перекрестноопыляемых растений. Пыльники во многих случаях растрескивались, но не у всех колосьев даже в пределах одного растения. У некоторых растений цветение было как бы смешанным, т. е. закрытым, если оно протекало при менее жаркой и сухой погоде или после полива, и открытым с нерастрескивающи-мися пыльниками у тех колосьев, мейоз, гаметогенез и цветение у которых совпало с особенно сухим и жарким периодом. Определение процентного содержания нормальной пыльцы в пыльниках у некоторых растений варьировало в очень широких пределах — от 10 до 70 и даже 80%.[ ...]
Как мы уже отмечали, в соответствии с заключением Т. Моргана гены сцеплены тогда, когда локализованы на одной хромосомной паре. Допуская, что хромосомы остаются интактными при вступлении их в гаметы, локализованные на них гены всегда наследуются вместе. В этом случае можно говорить об их полном сцеплении. Однако, рассматривая сцепление генов у кукурузы, мы видели, что полного сцепления генов не бывает, поскольку происходит формирование гамет не только двух родительских типов, но и гамет рекомбинантных типов (вследствие рекомбинации генов). Об отсутствии полного сцепления свидетельствуют также данные, полученные при изучении других организмов, механизм этого явления заключается в том, что в процессе гаметогенеза хромосомы клеток могут подвергаться разрывам в одном или нескольких местах, а сегменты, образующиеся в результате разрыва одной хромосомы, могут смыкаться с сегментами гомологичной хромосомы при условии, что в последней тоже были разрывы, причем в аналогичных местах. Как мы видели, гены С и в у кукурузы в 97% случаев (гамет) остаются сцепленными в родительских комбинациях и примерно в 3% случаев (гамет) они не связаны между собой и находятся в рекомбинантных сочетаниях. Обмен между хромосомными сегментами гомологичных хромосом, сопровождаемый рекомбинацией сцепленных генов, получил название кроссинговера (Т. Морган), а явление, обусловленное этим механизмом, называют генетической рекомбинацией. В результате рекомбинации из двух исходных комбинаций генов создается новая комбинация. Возвращаясь к случаю рекомбинации генов у кукурузы, можно сказать, что кроссинговер произошел в сегменте хромосомы между локуса-ми (местами), занимаемыми генами Сив, вследствие двух разрывов в этих участках хромосомы у отдельных клеток.[ ...]
Действительно, надо уточнять и упрощать шкалы для округления степени зрелости половых клеток и гонад у рыб, но во всех случаях указывать, какие критерии были взяты за основу при разграничении одной стадии зрелости от другой. С необходимостью уточнения универсальной шестибалльной шкалы зрелости мы столкнулись при анализе степени зрелости половых желез у морских тропических видов рыб, у самок которых чрезвычайно мелкая икра, основная оценка в большинстве случаев степени зрелости гонад осуществляется по объему яичника и его цвету, которые являются ■весьма изменчивым признаком. Однако, как показывают наши наблюдения совместно с кубинскими коллегами, сам тип развития ооцитов и процесс их выметывания совершенно иной по сравнению с гаметогенезом и экологией нереста наших пресноводных видов рыб средних широт.[ ...]
При анализе особенностей прохождения ежегодно повторяющихся половых циклов в водоемах различных широт было обнаружено, что в южных водоемах, в частности Куре и Кубани, самки судака зимуют с незрелыми половыми клетками. Процесс интенсивного вителлогенеза у них, в отличие от судака, обитающего в более северных водоемах, протекает незадолго до периода размножения судака в водоеме. Анализ полового цикла самок курин-ского судака показал, что особи в этом водоеме зимуют с половыми железами в III стадии зрелости яичников. На гистологических срезах яичников судака из р. Куры, которые были получены в ноябре, можно было видеть, что основная масса ооцитов в яйцене-сущих пластинках вступает лишь в начальный период трофоплаз-матического роста и находится в различных фазах вакуолизации и, в частности, в конечной фазе этого процесса, и лишь некоторые яйцеклетки в самом начале отложения питательных веществ (рис. 31, Б). В преднерестовый период ооциты генерации текущего года находятся в однородных фазах зрелости (рис. 31, А). При этом, так же как и у других исследованных нами представителей окуневых, у судака происходят изменения в строении и расположении запасных питательных веществ в ооцитах, обнаружены изменения в оболочках, ядре и т, д. В частности, по мере развития ооцита, происходит слияние мелких жировых вакуолей в более крупные, а затем образуется одна жировая капля, расположенная ближе к анимальному полюсу яйцеклетки. Ядро ооцита перед овуляцией смещается к вегетативному полюсу. Желток образует сплошную гомогенную массу. Многие из перечисленных особенностей развития яйцеклеток являются характерными для всего семейства в целом и подробно были описаны нами на примере развития половых клеток у окуня, как представителя семейства с наиболее четкими и последовательными картинами всего прохождения гаметогенеза и половых циклов.[ ...]